
الطفرة النقطية المقبولة (بالإنجليزية: Point accepted mutation) والمعروفة أيضًا باسم PAMهي تلك الطفرة التي يستبدل فيها حمض أميني واحد في التركيب الأساسي للبروتين بحمض أميني آخر، وهو ما يعد أمر مقبول خلال عمليات الاصطفاء الطبيعي. ولا يشمل هذا التعريف جميع أنواع الطفرات النقطية (Point mutations) في الحمض النووي للكائن الحي؛ حيث لا تعد الطفرات الصامتة أو الطفرات المميتة أو غيرها من الطفرات المرفوضة بواسطة الاصطفاء الطبيعي، من أنواع PAM.
هذا ويمثل كل عامود وصف في مصفوفة PAMاحد الأحماض الأمينية العشرين الأساسية. ولهذا تستخدم مصفوفات PAM، في المعلوماتية الحيوية، بانتظام كمصفوفات بديلة لتسجيل مدى انتظام تسلسل البروتينات. ويشير كل إدخال في مصفوفة PAM إلى احتمال استبدال الحمض الأميني في ذلك الصف بالحمض الأميني في ذلك العمود من خلال سلسلة واحدة أو أكثر من PAMخلال فترة تطور محددة، بدلاً من افتراض تراص هذين الحمضين الأمينين من باب الصدفة. بالإضافة إلى ذلك فأن مصفوفات PAM المختلفة تتوافق مع أطوال زمنية مختلفة في تطور تسلسل البروتين.
الخلفية البيولوجية
توجد التعليمات الوراثية لكل خلية مكررة في الكائن الحي، في الحمض النووي الخاص فيه [1]. تنسخ هذه المعلومات وتكرر، طيلة فترة حياة الخلية، بواسطة الآليات الخلوية لإنتاج البروتينات أو لتوفير إرشادات للخلايا الابنة أثناء انقسام الخلايا، وهنا يتواجد احتمال تغيير الحمض النووي خلال هذه العمليات [1] [2] والذي يؤدي إلى ما يعرف بالطفرة. أما على المستوى الجزيئي، فهناك أنظمة تنظيمية تصحح معظم - وليس كل - هذه التغييرات على الحمض النووي قبل تكرارها[2] [3].
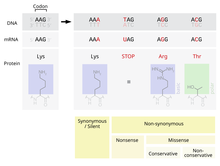
أحد الطفرات المحتملة التي يمكن أن تحدث هو استبدال نيوكلوتيدة واحدة أو زوج منها مما يؤدي إلى ما يعرف باسم الطفرة النقطية. وإذا ما حدثت الطفرة النقطية داخل الإكسون، فأن ذلك سيؤدي إلى تغيير الكودون الذي يحدد حمض أميني معين في البروتين الذي تنتجه تلك الجينة. وعلى الرغم من التكرار في الكود الوراثي، فأن هناك احتمال أن هذا الطفرة سوف تغير الأحماض الأمينية التي يتم إنتاجها أثناء عملية التشفير، ونتيجة لذلك سيتم تغيير بنية البروتين.
تعتمد وظيفة البروتين بشكل كبير على تركيبته [4] وقد يؤدي تغيير حمض أميني واحد في أحد البروتينات إلى التقليل من قدرته على تنفيذ هذه الوظيفة، أوقد يغير الوظيفة التي يؤديها البروتين [2]. مثل هذه التغييرات قد تؤثر بشدة على وظيفة حساسة في الخلية، مما قد يؤدي إلى موت الخلية أو حتى موت الكائن الحي في الحالات القصوى [5].من جهة أخرى، فإن هذا التغيير قد يسمح للخلية بمواصلة عملها وإن كان بشكل مختلف. علاوة على ذلك، يمكن أن تنتقل الطفرة إلى نسل الكائن الحي وفي حال عدم تسبب هذا التغيير بأي ضرر جسدي كبير للنسل، فاحتمالية استمرار هذه الطفرة في أوساط السكان كبيرة. ولا يمكن أن نغفل عن احتمالية أن يكون هذا التغييرفي الوظيفة مفيدًا. في كلتا الحالتين، فأن الطفرة النقطية قد قبلت في المجموعة الوراثية أثناء تعرضها لعمليات الاصطفاء الطبيعي.
على الرغم من تباين الأحماض الأمينية العشرين المشفرة بواسطة الشفرة الوراثية تباينًا كبيرًا وفقاًللخصائص الفيزيائية والكيميائية لسلاسلها الجانبية [4] إلا أنه يمكن تصنيف هذه الأحماض الأمينية إلى مجموعات ذات خصائص فيزيائية كيميائية متماثلة [4]. ومن المرجح أن يكون لاستبدال حمض أميني بحمض آخر من نفس الفئة تأثير أصغر على بنية البروتين ووظيفته من ما لو استبدل بحمض أميني من فئة مختلفة. وبالتالي، يعتمد قبول الطفرات النقطية اعتمادًا كبيرًا على الحمض الأميني الذي يتم استبداله في الطفرة وعلى بديل ذلك الحمض الأميني. هذا وتعد مصفوفات PAM أداة رياضية تراعي معدلات القبول المتفاوتة هذه عند تقييم تشابه البروتينات أثناء ترتيبها.
المصطلح
استخدم مصطلح الطفرة النقطية المقبولة (accepted point mutation) في البداية لوصف ظاهرة الطفرة، وفضل استخدام الاختصارPAM للتعبير عنه بدلاً من APM لأسباب تتعلق بمدى سهولة قراءته، لذلك يستعاض عن التسمية الأولى للمصطلح غالباً ب (point accepted mutation) [6].نظرًا لأن القيمة n في مصفوفة PAMnتمثل عدد الطفرات لكل 100 من الأحماض الأمينية، وبالتالي يمكن تسميتها بالنسبة المئوية للطفرات. نتيجة لذلك يمكن ملاحظة استخدام مصطلح النسبة المئوية للطفرة المقبولة أحيانًا
من المهم التمييز بين الطفرات النقطية المقبولة (PAMs) ومصفوفات الطفرة النقطية المقبولة (مصفوفاتPAM) ومصفوفة PAMn. حيث يشير مصطلح «الطفرة النقطية المقبولة» إلى حدث الطفرة نفسه. ويشير مصطلح «مصفوفة PAM» إلى مجموعة من المصفوفات التي تحتوي على درجات تمثل احتمالية محاذاة اثنين من الأحماض الأمينية بسبب سلسلة من أحداث الطفرة وليس بسبب الصدفة العشوائية. في حين أن «مصفوفةPAMn» هي مصفوفة PAM تتوافق وإطار زمني طويل بما يكفي لإحداث عدد (n)من الطفرات لكل 100 من الأحماض الأمينية.
بناء مصفوفات PAM
أول من قدم فكرة مصفوفات PAMهي مارجريت دايهوف Margare Dayhoff في عام 1978 [7]. واستند حساب هذه المصفوفات إلى 1572 طفرة ملحوظة في أشجار تطور السلالات، أخذت من 71 عائلة من البروتينات وثيقة الصلة. وتم اختيار البروتينات المراد دراستها بناءاً على وجود تشابه كبير مع سابقاتها. وكانت محاذاة البروتين مطلوبة لعرض تطابق ما لا يقل عن 85 ٪ [6] [8]. نتيجةً لذلك، من المنطقي افتراض أن عدم تطابق أي محاذاة هو نتيجة لحدث طفرة واحدة، بدلاً من افترض حدوثه نتيجة للعديد من الطفرات في نفس الموقع.
تحتوي كل مصفوفة من مصفوفات PAMعلى عشرين صفًا وعشرين عمودًا– كل منها تمثل الأحماض الأمينية العشرين المشفرة بواسطة الشفرة الوراثية. ترتبط القيمة في كل خلية من مصفوفة PAM باحتمال وجود صف من الحمض الأميني قبل محاذاة الطفرة مع عمود من الأحماض الأمينية بعد الطفرة [6] [7] [8].يمكن أن نستنتج من هذا التعريف بأن مصفوفاتPAM هي مثالاً لمصفوفة الاستبدال.
جميع البيانات من اشجار تطور السلالات
سجل لكل فرع من فروع أشجار تطور السلالات لعائلات البروتين عدد من حالات عدم التطابق التي تمت ملاحظتها بالإضافة إلى تسجيل الحمضين الأمينيين المعنيين بذلك [7]. واستخدمت هذه التعدادات كإدخالات أسفل القطر الرئيسي للمصفوفة A. ونظرًا لأن الغالبية العظمى من عينات البروتين تأتي من كائنات حية اليوم، فأنه من الصعب تحديد «اتجاه» الطفرة. أي أن الحمض الأميني الموجود قبل الطفرة لايمكن تمييزه عن الحمض الأميني الذي حل محله بعد الطفرة. لهذا السبب، يفترض أن المصفوفة A متناظرة، ويتم حساب إدخالات A أعلى القطر الرئيس على هذا الأساس. الإدخالات على طول قطري A لا تتوافق مع الطفرات ويمكن تركها بدون تعبئة.
بالإضافة إلى هذه الإحصائيات، تم الحصول على بيانات حول قابلية التحول وتواتر الأحماض الأمينية[6] [7]. حيث أن قابلية تحول الأحماض الأمينية هي نسبة عدد الطفرات التي تشارك فيها وعدد مرات حدوثها في المحاذاة. [7] وتقيس قابلية التحول مدى احتمال تحول الحمض الأميني إلى طفرة. ووجد أن الأسباراجين، وهو حمض أميني ذي سلسلة جانبية قطبية صغيرة، ليكون أكثر الأحماض قابلية للتحول من بين الأحماض الأمينية. [7] في حين أن كل من السيستين والتريبتوفان هما الأقل قابلية للتحول من بين الأحماض الأمينية. [7] حيث تحتوي السلاسل الجانبية للسيستين والتريبتوفان على هياكل أقل شيوعًا؛ فتحتوي السلسلة الجانبية لسيستين على الكبريت الذي يشارك في روابط ثاني كبريتيد مع جزيئات السيستين الأخرى، أما التريبتوفان فيحتوي على أكبر سلسلة جانبية عطرية بين الأحماض الأمينية. [4] نظرًا لوجود العديد من الأحماض الأمينية القطبية الصغيرة، فإن هذه الحالات المتطرفة تشير إلى أن الأحماض الأمينية أكثر عرضة للطفرة إذا كانت خواصها الفيزيائية والكيميائية أكثر شيوعًا بين الأحماض الأمينية البديلة. [6] [8]
بناء مصفوفات الطفرة
في ما يتعلق بالحمض الأميني jth فأن قيمة و تمثلان قيمت التحول والتواتر. وتضبط تواترات الأحماض الأمينية بحيث تصل إلى 1. إذا كان إجمالي عدد مرات حدوث الحمض الأميني jth هو و N هي المجموع الكلي للأحماض الأمينية، إذن:

وبناءً على تعريف قابلية التحول (التطفر) كنسبة الطفرات إلى ظهور الحمض الأميني، فأن

أو

يتم إنشاء مصفوفة الطفرة M بحيث يمثل الإدخال احتمال تحول الحمض الأميني jth إلى الحمض الأميني ith. يتم حساب الإدخالات غير القطرية بواسطة المعادلة التالية: [7]

حيث يمثل ثابت التناسب. ومع ذلك، فإن هذه المعادلة لا تحسب الإدخالات القطرية. حيث يسرد كل عامود في المصفوفة M النتائج العشرين المحتملة للحمض الأميني–وإذا ما يمكن أن يتحول إلى واحد من التسعة عشر حمض أميني أخر، أو يبقى على ما هو عليه. نظرًا لأن الإدخالات غير القطرية تسرد احتمالات كل من الطفرات التسعة عشر المعروفة، والتي يجب أن يكون مجموع احتمالات هذه النتائج العشرين 1، يمكن حساب هذا الاحتمال الأخير بواسطة المعادلة التالية:

والتي يمكن تبسيطها إلى:

نتوصل لنتيجة ذات أهمية كبيرة وهي أن للإدخالات غير القطرية يمكن اتباع المعادلة التالية:

والتي تعني أنه بالنسبة لجميع الإدخالات في مصفوفة الطفرة

اختيار ثابت التناسب
تختلف الاحتمالات الموجودة في M نتيجة لعدم معرفة مقدار الوقت المسموح فيه لتسلسل البروتين ليتحول (يطفر) فيه. بدلاً من محاولة تحديد هذه العلاقة، يتم حساب قيمM في إطار زمني قصير، ويتم حساب المصفوفات لفترات زمنية أطول من خلال افتراض أن الطفرات تتبع نموذج سلسلة ماركوف. [9] [10] الوحدة الأساسية للوقت لمصفوفات PAM هي الوقت اللازم لحدوث طفرة واحدة لكل 100 من الأحماض الأمينية، والتي تسمى أحيانًا «وحدة PAM» أو "PAM" من الوقت. [6] هذه هي مدة الطفرة التي تفترضها مصفوفة PAM1 بالضبط.
يستخدم الثابت للتحكم في نسبة الأحماض الأمينية التي لم تتغير. باستخدام محاذاة البروتينات التي تتشابه بمقدار 85٪ على الأقل، يمكن افتراض أن الطفرات التي لوحظت كانت مباشرة، دون أي حالات وسيطة. هذا يعني أن خفض هذه الإحصاءات بعامل مشترك من شأنه أن يوفر تقديراً دقيقاً لتعداد الطفرات لو كان التشابه أقرب إلى 100٪. وهذا يعني أيضًا أن عدد الطفرات لكل 100 من الأحماض الأمينية حيث n في PAMn يساوي عدد الأحماض الأمينية المتحولة لكل 100 من الأحماض الأمينية.
للعثور على مصفوفة الطفرة في مصفوفة PAM1 ، يتم فرض متطلبات حفظ 99 ٪ من الأحماض الأمينية في التسلسل. الكمية n(j)M(j,j) تساوي عدد وحدات الأحماض الأمينية المحفوظة، وبالتالي فإن إجمالي عدد الأحماض الأمينية المحفوظة هو:

ثم يتم إعطاء قيمة اللازمة لإنتاج 99٪ تطابق بعد حدوث طفرة بواسطة المعادلة

يمكن عندئذ استخدام القيمة في مصفوفة الطفرة لمصفوفة PAM1
بناء مصفوفات PAM n
نموذج سلسلة ماركوف من طفرة البروتين يرتبط بمصفوفة التحوّل لـ PAMn، Mn ، بمصفوفة التحوّل في مصفوفة، بواسطة العلاقة البسيطة

يتم إنشاء مصفوفة PAMn من نسبة احتمال حدوث الطفرات النقطية المقبولة التي تستبدل الحمض الأميني jth بالحمض الأميني ith ، إلى نسبة احتمال أن يكون تحاذي هذه الأحماض الأمينية عن طريق الصدفة. يتم إعطاء إدخالات مصفوفة PAMnوفق المعادلة التالية [11] ]12[

لاحظ أنه في كتاب جسفيلد، ترتبط المدخلات و باحتمال تحوّل الحمض الأميني ith إلى الحمض الأميني 11] jth ]. وهذا ما يعد أصل المعادلة المختلفة لإدخالات مصفوفات PAM.
عند استخدام مصفوفة لتسجيل محاذاة بروتينين، يتم الافتراض التالي:
- إذا كان هذان البروتينين مرتبطين، فإن الفاصل التطوري الذي يفصل بينهما هو الوقت الذي يستغرقه حدوث n من الطفرات النقطية المقبولة لكل 100 من الأحماض الأمينية.
عند النظر في محاذاة الأحماض الأمينية ith و jth ، فإن النتيجة تشير إلى الاحتمالات النسبية للمحاذاة بسبب ارتباط البروتينات أو بسبب فرصة عشوائية.
- · إذا كانت البروتينات مرتبطة، يجب أن تكون هناك سلسلة من الطفرات النقطية المقبولة قد حدثت لتحويل الحمض الأميني الأصلي إلى بدائله. وعلى فرض أن الحمض الأميني jth هو الأصلي، واستنادًا إلى وفرة الأحماض الأمينية في البروتينات، يكون احتمال أن يكون الحمض الأميني jth الأصلي هو f(j). بالنظر إلى أي وحدة من هذا الحمض الأميني، فإن احتمال استبداله بحمض أميني ith في الفاصل الزمني المفترض هو . وبالتالي، فإن احتمال المحاذاة هو، وهو البسط داخل اللوغاريتم. · إذا لم تكن البروتينات مرتبطة، فإن الأحداث التي يكون فيها الأحماض الأمينية المتحاذيان هما ith و jth يجب أن تكون مستقلة. احتمالات هذه الأحداث هي، مما يعني أن احتمال المحاذاة هو، مقام اللوغاريتم. · وبالتالي، ينتج عن اللوغاريتم في المعادلة إدخالًا إيجابيًا إذا كان حدوث المحاذاة بسبب الطفرات النقطية المقبولة هو الأكثر ترجيحًا، وإدخالًا سلبيًا إذا كان حدوث المحاذاة نتيجة للصدفة هو الأكثر ترجيحًا.
خصائص مصفوفات PAM
التماثل في مصفوفات PAM
على الرغم من أن مصفوفة احتمال الطفرة M ليست متماثلة، فإن كل مصفوفة من مصفوفات PAM متماثلة. [6] [7] هذه الخاصية المدهشة إلى حد ما هي نتيجة العلاقة التي تمت الإشارة إليها في مصفوفة احتمالية حدوث الطفرة:
في الحقيقة، تحمل هذه العلاقة جميع القوى الصحيحة الموجبة للمصفوفة
ونتيجة لذلك، فإن إدخالات مصفوفة PAMn متماثلة؛ بما أن


يربط عدد الأحماض الأمينية المتحولة وعدد الطفرات
تمثل القيمة n عدد الطفرات التي تحدث لكل 100 من الأحماض الأمينية، ومع ذلك، نادراً ما تكون هذه القيمة متاحة وغالبًا ما يتم تقديرها. ومع ذلك، عند مقارنة بروتينين، من السهل حساب m بدلاً من ذلك، وهو عدد الأحماض الأمينية المتحولة لكل 100 من الأحماض الأمينية. على الرغم من الطبيعة العشوائية للطفرة، يمكن ربط هذه القيم تقريبًا بواسطة المعادلة التالية[13]

يمكن التحقق من صحة هذه التقديرات من خلال حساب عدد الأحماض الأمينية التي لا تزال دون تغيير في إطار عمل المصفوفة M. العدد الإجمالي للأحماض الأمينية غير المتغيرة للفترة الزمنية لمصفوفة PAMn، هو:

وبالتالي فإن نسبة الأحماض الأمينية التي لم تتغير، هي:

مثال - PAM250
PAM 250 هي مصفوفة تسجيل شائعة الاستخدام لمقارنة التسلسل، وفيها يجب حساب النصف السفلي فقط من المصفوفة. حيث إنه من خلال بنائها، يجب أن تكون مصفوفات PAM متماثلة. يظهر كل من الأحماض الأمينية العشرين أسفل الجزء العلوي من المصفوفة وجانبها، مع 3 أحماض أمينيه غامضة إضافية. تظهر الأحماض الأمينية في الغالب مدرجة في قائمة أبجدية أو مدرجة في مجموعات. هذه المجموعات هي الخصائص المشتركة بين الأحماض الأمينية. [7]

الاستخدم في مجال المعلوماتية الحيوية
تحديد وقت الاختلاف في اشجار تطور السلالات
تتنبأ فرضية الساعة الجزيئية بأن معدل إحلال الأحماض الأمينية في بروتين معين سيكون ثابتًا تقريبًا بمرور الوقت، على الرغم من أن هذا المعدل قد يختلف بين عائلات البروتين. [13] هذا يشير إلى أن عدد الطفرات في الأحماض الأمينية في البروتين يزداد خطيًا تقريبًا مع مرور الوقت.
إن تحديد الوقت الذي ينفصل فيه بروتينان مُهِمةٌ مهمة في علم الوراثة. غالبًا ما تستخدم السجلات الأحفورية لتحديد موقع الأحداث على الجدول الزمني للتاريخ التطوري للأرض، ولكن تطبيق هذا المصدر محدود. ومع ذلك، إذا كان المعدل الذي تحدده الساعة الجزيئية لعائلة البروتين - أي المعدل الذي يزداد فيه عدد الطفرات في الأحماض الأمينية - معروفًا، فإن معرفة هذا العدد من الطفرات سيتيح تحديد تاريخ الانفصال.
لنفترض بأننا نبحث في تاريخ الانفصال بين بروتينين مرتبطين، مأخوذين من الكائنات الحية التي تعيش اليوم . حيث قام كلا البروتينين بتجميع طفرات مقبولة منذ تاريخ الانفصال، وبالتالي فإن العدد الإجمالي للطفرات لكل حمض أميني يفصل بينهما هو تقريبا ضعف العدد الذي يفصلهما عن سلفهما المشترك. وإذا استخدمت مجموعة من مصفوفات PAM لمحاذاة بروتينين معروفين بكونهما مرتبطان، فمن المرجح أن تتطابق قيمة n في مصفوفة PAMn، والذي بدوره سيؤدي إلى أفضل درجة، مع الطفرات في الحمض الأميني الذي يفصل بين البروتينين. يوفر خفض هذه القيمة وتقسيمها على المعدل الذي تتراكم فيه الطفرات المقبولة في عائلة البروتين، تقديراً لوقت انفصال هذين البروتينين عن سلفهما المشترك. هذا هو، وقت الانفصال في myr :[13]

حيث K هو عدد الطفرات لكل حمض أميني، و r هو معدل تراكم الطفرة المقبول في الطفرات في موقع الحمض الأميني لكل مليون سنة.
الاستخدام في الBLAST
يتم أيضًا استخدام مصفوفات PAM كمصفوفة تسجيل عند مقارنة تسلسل الحمض النووي أو تسلسل البروتين للحكم على جودة المحاذاة. يتم استخدام هذا النوع من نظام التسجيل من خلال مجموعة واسعة من برامج المحاذاة بما في ذلك BLAST.
مقارنة بين PAM و BLOSUM
بالإضافة إلى مصفوفات PAM ، يمكن استخدام مصفوفة تسجيل أخرى تم تطويرها مؤخرًا والتي تعرف باسم BLOSUM. ينتج الاثنان نفس نتيجة التسجيل، لكنهما يستخدمان منهجيات مختلفة. في حين تنظر BLOSUM مباشرةً إلى الطفرات في أشكال التسلسلات ذات الصلة، تقوم PAM باستقراء المعلومات التطورية استنادًا إلى متواليات وثيقة الصلة. [16]
ونظرًا لأن كلاً من PAM و BLOSUM هما طريقتان مختلفتان لإظهار معلومات التسجيل نفسها، فيمكن مقارنة الاثنين ولكن وبسبب الطريقة المختلفة جدًا للحصول على هذه النتيجة، فإن PAM100 لا تساوي BLOSUM100 . [17]
انظر أيضا
- طفرة نقطة
- محاذاة التسلسل
- مارغريت دايهوف
- الساعة الجزيئية
- BLOSUM
- انفجار