
|
فيروس كورونا | |
|---|---|
 | |

تمثيل لفيروس سارس-كوف-2 الذي سبب جائحة كوفيد 19.
الأزرق: غلاف ليبيد ثنائي الطبقة أزرق فاتح: بروتين الشوكة السكري أحمر: بروتين الغلاف أخضر: بروتين الغشاء برتقالي: جليكان | |
| المرتبة التصنيفية | أسرة |
| تصنيف الفيروسات | |
| المجموعة: | فيروس |
| النطاق: | ريبوفيريا |
| المملكة: | أرثوورنافيرات |
| الشعبة: | بيسوفيريكوتا |
| الطائفة: | بيسونيفريسيتس |
| الرتبة: | فيروسات عشية |
| الرتيبة: | كورنيدوفرينيات |
| الفصيلة: | فيروسات تاجية |
| الأسرة: | فيروس كورونا |
| الاسم العلمي | |
| Orthocoronavirinae | |
| الأجناس | |
| مرادفات | |
| كورونافيرينات | |
|
تعديل مصدري - تعديل | |
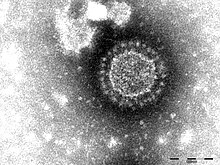
فيروسات كورونا أو الكورونافيرينات المستقيمة (الاسم العلمي: Orthocoronavirinae) هي عائلة فرعية من فيروسات الرنا مفردة السلسلة موجبة الدلالة المغلفة التي تصيب الثدييات والطيور، وتنتمي إلى عائلة الفيروسات التاجية من رتبة الفيروسات العشية. أول فيروس كورونا تم تحديده هو فيروس التهاب الشعب الهوائية المعدي الذي يصيب الدجاج في عقد 1920، واكتُشفت فيروسات كورونا التي تسبب الزكام للبشر في عقد 1960. سميت هذه الفيروسات باسم كورونا بسبب مظهرها تحت المجهر الإلكتروني الذي يبدو كهالة أو تاج بسبب بروتينات الشوكة.
يتراوح طول جينوم فيروسات كورونا بين 26 ألف و32 ألف نوكليوتيد وهو أطول الجينومات بين فيروسات الرنا، ويشفر أربع بروتينات بنيوية: الشوكة (S)، الغلاف (E)، الغشاء (M)، القفيصة النووية (N) وفي بعض الفيروسات بروتين خامس هو إستراز الراصة الدموية (HE) كما يشفر جين الريبلكاز والعديد من البروتينات الملحقة. فضلا على الحفاظ على بنية الفيروس، تملك هذه البروتينات وظائف متعددة مثل تعزيز الإصابة ومقاومة الاستجابات المناعية للمضيف. يقوم بروتين الشوكة بالارتباط بمستقبلاتٍ على سطح الخلية المضيفة ودمج الغلاف الفيروسي مع الغشاء الخلوي الخاص بالخلية المضيفة للدخول إليها. بعد الدخول إلى السيتوبلازم تعمل بعض البروتينات اللابنيوية على إعادة تنظيم الغشاء الداخلي لإنشاء عضيات التضاعف التي يحدث فيها النسخ والتضاعف. أثناء النسخ، يحدث تبديل للقالب لإنشاء جزيئات رنا جينومية فرعية بأطوال مختلفة تشفر واحدا أو عدة بروتينات بنيوية وهي ميزة تتميز بها الفيروسات العشية، قد يكون تبديل القالب هو آلية التأشيب الجيني بين مختلف فيروسات كورونا التي تسمح لها بتوليد سلالات جديدة أو الانتقال عبر الأنواع. يخضع جين الريبليكاز لانزياح إطار ريبوسومي -1 أثناء الترجمة لنسخ عديد البروتين الكبير 1ab الذي يخضع لقص ذاتي بواسطة وظيفة البروتياز الخاصة بالبروتينين اللابنيويين 3 و5 وينتج عنه 16 بروتينا لا بنيويا لها وظائف متنوعة أهمها البروتين اللابنيوي 12 الذي يملك وظيفة بوليميراز الرنا المعتمد على الرنا ويقوم بعملية النسخ، والبروتين اللابنيوي 14 الذي يملك وظيفة تصحيح تزيد من دقة النسخ، وربما يكون السبب في قدرة فيروسات كورونا على امتلاك جينوم بهذا الطول. الإصابة بفيروس كورونا تسبب استجابات خلوية عديدة مثل استجابة إجهاد الشبكية الإندوبلازمية التي تؤدي إلى إفراز السيتوكينات وحدوث الاستماتة، وتؤثر على العديد من مسارات التأشير وتثير استجابات مناعية، لكن يملك الفيروس بعض الآليات لمقاومة هذه الاستجابات وتثبيط مسارات تخليق الإنترفيرونات.
تنقسم عائلة فيروسات كورونا الفرعية إلى أربع أجناس: فيروس كورونا ألفا، فيروس كورونا بيتا، فيروس كورونا غاما، فيروس كورونا دلتا وتصيب أجناس ألفا وبيتا الثدييات فقط وسلفهما المشترك يمكن أن يكون فيروسا خفاشيا، أما غاما ودلتا فيصيب معظمها الطيور بشكل رئيسي مع أنواع قليلة تصيب الثدييات ويُفترض أن سلفهما المشترك فيروس يصيب الطيور. ينتمي فيروسا سارس-كوف وسارس-كوف-2 إلى نوع الفيروسات المرتبطة بالسارس الذي ينتمي إلى الجنس الفرعي الفيروسات الساربيكوية الذي ينتمي إلى جنس فيروسات بيتا. يشمل الجنس الفرعي فيروس إمبيكو الفيروسات التي تصيب الفئران ومن بينها فيروس التهاب الكبد الفأري وهو فيروس نموذجي في الأبحاث وقد تم توضيح العديد من الآليات الجزيئة لفيروسات كورونا عبر دراسته.
توجد سبع فيروسات تصيب البشر، 4 منها (فيروس كورونا البشري OC43، فيروس كورونا البشري HKU1، فيروس كورونا البشري 229E وفيروس كورونا البشري NL63) يمكن أن تسبب الزكام، والثلاثة الأخرى سببت تفشيات أمراض خطيرة حول العالم وهي فيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة النوع 1 (سارس-كوف)، فيروس كورونا المرتبط بمتلازمة الشرق الأوسط التنفسية (ميرس-كوف) وفيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة النوع 2 (سارس-كوف-2). فضلا عن ذلك، توجد العديد من فيروسات كورونا التي يمكن أن تصيب الماشية والدواجن والحيوانات الأليفة وحيوانات المختبر والحيوانات البرية، مثل فيروس كورونا الطيري الذي يصيب الدجاج والعديد من فيروسات كورونا الخنزيرية التي تصيب الخنازير وفيروس كورونا الكلبي وفيروس كورونا القططي اللذان يصيبان الكلاب والقطط، وفيروس كورونا مورين الذي يصيب الفئران والجرذان المختبرية.
التأثيل
اشتُق اسم "coronavirus" (عربيًا: فيروس كورونا. اختصارًا CoV) من الكلمة (اللاتينية corona) وتعني «التاج» أو «الإكليل» المأخوذة بدورها من الكلمة (الإغريقية κορώνη) وتعني إكليل زهور أو الإكليل. صيغ المصطلح بواسطة جون ألميدا وديفيد تيريل اللذان كانا أول من لاحظ ودرس فيروسات كورونا البشرية. استُعمل المصطلح لأول مرة في منشور سنة 1968 بواسطة مجموعة من علماء الفيروسات غير الرسميين في مجلة نيتشر لتسمية عائلة الفيروسات الجديدة. يُشير الاسم إلى المظهر المميز للفيريونات (الشكل المُعدي للفيروس) والذي يظهر عبر المجهر الإلكتروني، حيث تمتلك خُملًا/زغاباتٍ من البروزات السطحية البصلية الكبيرة، مما يُظهرها على شكل تاج الملك أو الهالة الشمسية. يحدث هذا التشكُل عبر قسيماتٍ فولفية للشوكة الفيروسية (S)، وهي بروتيناتٌ تملأ سطح الفيروس وتحدد انتحاء المضيف.
قُبل الاسم العلمي "Coronavirus" بواسطة اللجنة الدولية لتسمية الفيروسات التي أعيدت تسميتها لاحقا إلى اللجنة الدولية لتصنيف الفيروسات (ICTV) في عام 1971. ومع ازدياد عدد الأنواع تم تقسيم هذه العائلة الفرعية إلى أربع الجنس هي: فيروس كورونا ألفا، فيروس كورونا بيتا، فيروس كورونا غاما وفيروس كورونا دلتا عام 2009. يُستخدم الاسم الشائع فيروس كورونا للإشارة إلى أي عضو في العائلة الفرعية "Orthocoronavirinae" من عائلة الفيروسات التاجية. واعتبارا من 2020، يوجد 45 نوعا من فيروسات كورونا معترف بها رسميا.
في اللغة العربية، تُعتبر تسمية فيروس كورونا(1) أكثر شيوعًا من باقي التسميات الأُخرى، ولكن التسميات الأُخرى أكثر دقةٍ في الوصف، حيثُ يُسمى: الفيروس التاجي، فيروس الهالة، الفيروسة المُكَلَّلَة (أو الفيروس المكلل)، الحُمّة التاجيّة الحُمَة الإكليلية، الحمة التاجية المكللة.
تاريخ الأبحاث

ظهرت أولى التقارير عن إصابة حيواناتٍ بفيروس كورونا في أواخر العقد 1920، عندما تفشت عدوى تنفسية حادة بين دجاجٍ مستأنس في أمريكا الشمالية. حيث كتب آرثر شالك وم.س. هاون عام 1931 أول تقرير مفصل يصف عدوى تنفسية جديدة بين الدجاج في داكوتا الشمالية. اتسمت إصابة الكتاكيت حديثة الولادة باللهاث والفتور مع معدلات نفوق عالية بين 40-90٪. عزل ليلاند ديفيد وكارل ألفريد الفيروس الذي تسبب في الإصابة بالعدوى عام 1933، وعُرف آنذاك باسم فيروس التهاب الشعب الهوائية المعدي (IBV). تمكن تشارلز د.هدسون وفريد روبرت بيوديت من زرع الفيروس لأول مرة عام 1937، وأصبحت تلك العينة تُعرف باسم سلالة بيوديت. في أواخر العقد 1940، اكتُشف نوعان آخران من فيروسات كورونا الحيوانية: JHM الذي يتسبب في مرض الدماغ (التهاب الدماغ عند الفئران) وفيروس التهاب الكبد الفأري (MHV) الذي يسبب التهاب الكبد في الفئران. لم يتم في ذلك الوقت إدراك أن هذه الفيروسات الثلاثة المختلفة كانت لها صلة ببعضها.
اكتُشفت فيروسات كورونا البشرية في العقد 1960 باستخدام طريقتين مختلفتين في المملكة المتحدة والولايات المتحدة. جمع إ.س. كيندال، ومالكولم بينو، وديفيد تيريل في عام 1961 أثناء عملهم في وحدة الزكام التابعة لمجلس البحوث الطبية البريطاني فيروسا فريدا يسبب الزكام سُمي بـB814. ولم يتمكنوا من زراعته باستخدام التقنيات المعيارية التي نجحت سابقا في زراعة الفيروسات الأنفية والفيروسات الغدية وغيرها من الفيروسات التي تسبب الزكام. في عام 1965، نجح تيريل وبينو في زراعة الفيروس الجديد عبر تمريره بشكل متسلسل في مزارع أعضاء لقصبات هوائية جنينية بشرية. وهي طريقة زراعة جديدة أحضرها برتيل هورن إلى ذلك المختبر. تسبَّبَ الفيروس المعزول عند تلقيح المتطوعين به عن طريق الأنف في الإصابة بالزكام وتم تثبيطه بواسطة الإيثر وهو ما يشير إلى امتلاكه لغلاف دهني. عزلت دوروثي هامري وجون بروكنو في جامعة شيكاغو فيروس زكام جديد من طلاب طبٍ في عام 1962، وقاما بتنميته في مزرعة نسيج الكلى وسمياه 229E. سبّب الفيروس الجديد زكاما لدى المتطوعين، وعلى غرار B814 ، تم تثبيطه بواسطة الإيثر.

قارنت عالمة الفيروسات الاسكتلندية جون ألميدا من مستشفى سانت توماس في لندن -بالتعاون مع تيريل- بُنى الفيروسات IBV وB814 و229E في عام 1967. باستخدام المجهر الإلكتروني، تبين أن الفيروسات الثلاثة لها صلة مورفولوجيا من خلال شكلها العام وشوكاتها المميزة التي تشبه النفل. تمكنت مجموعة بحثية في معاهد الصحة الوطنية الأمريكية في نفس العام من عزل عضو آخر من هذه المجموعة الجديدة من الفيروسات باستخدام زراعة الأعضاء وقامت بتسمية إحدى العينات OC43 (OC اختصارٌ لـorgan culture). على غرار B814 و229E وIBV، كان لفيروس الزكام الجديد OC43 شوكات مميزة شبيهة بالنفل عندما لوحظ بالمجهر الإلكتروني.
سرعان ما تبين أن فيروسات الزكام الجديدة المماثلة لـ IBV لها صلة مورفولوجية بفيروس التهاب الكبد الفأري، وسُميت هذه المجموعة الجديدة من الفيروسات باسم فيروسات كورونا وذلك لمظهرها المورفولوجي المميز. استمرت دراسة فيروس كورونا البشري 229E وفيروس كورونا البشري OC43 خلال العقود اللاحقة، أما سلالة فيروس كورونا B814 فقد فُقدت ومن غير المعروف أيٌّ من فيروسات كورونا الحالية هو فيروس B814. منذ ذلك الحين، تم تحديد عدة فيروسات كورونا بشرية أخرى ومنها: سارس-كوف في عام 2003، وكوف-NL63 البشري في عام 2003، وكوف-HKU1 البشري في عام 2004، وميرس-كوف في عام 2013، وسارس-كوف-2 في عام 2019. كما تم تحديد عدد كبير من فيروسات كورونا الحيوانية منذ الستينيات.
علم الفيروسات
الشكل والبنية

فيروسات كورونا هي جسيمات كبيرة، كروية الشكل تقريبا وذات نتوءات (شوكات) سطحية فريدة. حجمها متغير بدرجة كبيرة ومتوسط قطرها يترواح بين 80 إلى 120 نانومتر، ويصل قطر الأحجام المختلفة إلى حد أدناه 50 ناتومتر وأقصاه 200 نانومتر. يبلغ متوسط الكتلة الجزيئية الكلية 40000 كيلو دالتون. وهي محاطة بغلاف مدمجة فيه عدة جزيئات بروتينية. يحمي غلاف الليبيد ثنائي الطبقة وبروتينات الغشاء والقفيصة النووية الفيروس عندما يكون خارج الخلية المضيفة.
يتكون غلاف الفيروس من ليبيد ثنائي الطبقة تكون البروتينات البنيوية: الغشاء (M) والغلاف (E) والشوكة (S) مدمجة به. نسبة تواجد هذه البروتينات في غشاء اللبيد ثنائي الطبقة هي حوالي 1 بروتين غلاف لكل 20 بروتين شوكة لكل 300 بروتين غشاء. بروتينا الغلاف والغشاء هما المسؤولان على تحديد شكل الفيروس حيث يتآثران مع الليبيد ثنائي الطبقة لإحداث انحناءات في الغشاء وإنشاء فيروس شكله كروي تقريبا وتحديد حجمه. بروتين الشوكة ضروري من أجل التآثر مع مستقبلات على سطح الخلايا المضيفة من أجل الدخول، لكن فيروس كورونا البشري NL63 متميز بأن بروتين الغشاء هو من لديه نطاق الارتباط بمستقبلات الخلية المضيفة وليس بروتين الشوكة. يبلغ قطر الغلاف حوالي 85 نانومتر. يظهر غلاف الفيروس في الصور المجهرية الإلكترونية كزوج مميز من أغلفة كثافة الإلكترون (غلافان معتمان نسبيا مقارنة بشعاع الإلكترون المستخدم لمسح جسيم الفيروس).

الجينوم
تملك فيروسات كورونا جينوم رنا مفرد السلسلة موجب الدلالة، يتراوح طوله بين 26 ألف إلى 32 ألف نوكليوتيد. وهو من أكبر الجينومات بين فيروسات الرنا. يحتوي الجينوم على قبعة 5' ممثيلة في النهاية 5' وذيل عديد أدينين في النهاية 3' طوله بين 30-60 أدينين.
تنظيم الجينات في جينوم فيروسات كورونا عموما هو: المنطقة غير المترجمة 5' - الريبلكاز - الشوكة - الغلاف - الغشاء - القفيصة النووية - المنطقة غير مترجمة 3' - ذيل عديد الأدينين. يشغل إطارا القراءة المقتوحان 1a و1b ثلثي الجينوم ويشفران عديد بروتين الريبلكاز 1ab الذي يقوم بقص نفسه إلى 16 برووتينا لا بنيويا. تشفر إطارات القراءة المفتوحة التالية البروتينات البنيوية الرئيسية الأربعة: الشوكة والغلاف والغشاء والقفيصة النووية، وفي بعض الفيروسات يوجد بروتين بنيوي خامس هو إستراز الراصة الدموية (HE). تتخلل هذه البروتينات البنيوية بروتينات ملحقة يختلف عددها وموضعها ووظيفتها وطولها حسب الفيروس، ورغم أن معظمها غير لازم من أجل تضاعف الفيروس أو إصابته للخلية إلا أنها مسؤولة على الوظيفة الإمراضية للفيروس وتلعب أدوارا في مقاومة الاستجابة المناعية للمضيف.
المنطقة غير المترجمة 5' و3'
المنطقة غير المترجمة 5' واختصارا مغم 5' (5′ UTR) والمنطقة غير المترجمة 3' (مغم 3') هما منطقتان من جينوم فيروس كورونا لا تتم ترجمتهما إلى بروتينات لكنهما ضروريتان ولازمتان من أجل تنسخ الفيروس وتضاعفه، وتحتويان على العديد من الحلقات جذعية التي تسلسلاتها وبُناها الثانوية منحفظة تطوريًا.
المنطقة غير المترجمة 5'

يبلغ طول المنطقة غير المترجمة 5' الخاصة بفيروس كورونا حوالي 200-300 نوكليوتيد، وتحتوي بشكل عام على الحلقة الجذعية 1 (SL1) والحلقة الجذعية 2 (SL2) والحلقة الجذعية 4 (SL4)، ويملك فيروس كورونا البقري، وفيروس كورونا المرتبط بالسارس وبعض فيروسات كورونا غاما ودلتا أيضا حلقة جذعية 3 (SL3) بين الحلقة الجذعية 2 والحلقة الجذعية 4. فضلا عن هيئة التطوي الكاملة، يمكن أن تتواجد SL1 كذلك في توازن ديناميكي مع هيئات تطوي عالية الطاقة غير كاملة التطوي في الجذع، تعمل بمساعدة بروتينات مفترضة على التآثر مع مناطق أخرى من الجينوم على مسافات طويلة والمشاركة في تنسخ الفيروس وتضاعفه. تسلسل الحلقة SL2 هو التسلسل الأكثر انحفاظا بين تسلسلات الحلقات الجذعية في مغم 5'، ويمكن أيضًا أن يكون له صلة بتنسخ الفيروس وتضاعفه. تحتوي الحلقة SL3 على التسلسل المنظم للنسخ الطليعي (TRS-L) في منطقة الحلقة وهو أساسي وحاسم لتخليق الرنا الجينومي الفرعي أثناء تضاعف الفيروس، لكن هذه الحلقة غير مستقرة في فيروسات مثل فيروس كورونا الفأري. تسلسل الحلقة SL4 طويل نسبيًا ويمكن تقسيم الحلقة الجذعية إلى جزأين: 4a و4b مفصولين بانتباج في المنتصف. يوجد إطار قراءة مفتوح قصير في SL4 في العديد من فيروسات كورونا هو إطار القراءة المفتوح عكس الاتجاه، الذي يمكنه تثبيط ترجمة الريبليكاز 1ab، تُظهر النتائج التجريبية أن SL4 متسامحة للغاية مع الطفرات وتتحملها وحتى استبدال SL4 كاملة بتسلسل مختلف لا يؤثر على التضاعف (ولكن إذا تم حذفها لا يمكن أن يتضاعف الفيروس)، لذلك افتُرض أن وظيفتها الرئيسية هي تحديد الرنا الجينومي الفرعي الذي يتم نسخه، وبنية جذعها أكثر مرونة وهو ما يسمح لها بالتآثر مع الرنا الجينومي الفرعي أثناء النسخ والمساعدة في تبديل القالب.
علاوة على هذه الحلقات، تملك العديد من فيروسات كورونا حلقة خامسة SL5 مع الاتجاه بالنسبة للحلقة الرابعة. تتجاوز الحلقة 5 في فيروسات ألفا وبيتا المنطقة غير المترجمة 5' وتتداخل مع ORF1a وتسلسلها أكثر انحفاظا بين فيروسات ألفا. تتكون الحلقة 5 في فيروسات ألفا وبيتا من ثلاث حلقات صغيرة SL5a وSL5b وSL5c جميعها تحتوي على التسلسل 5'-UUCCG(U/C)-3 في أعلاها. إشارة التغليف ضرورية من أجل تجميع بعض فيروسات كورونا. تملك فيروسات غاما الحلقة SL5 كذلك لكن بنييتها مختلفة قليلا عن تلك الخاصة بفيروسات ألفا وبيتا، ويمكن أن يكون لها أيضا دور في تضاعف الرنا الفيروسي.
المنطقة غير المترجمة 3'

يبلغ طول المنطقة غير المترجمة 3' الخاصة بفيروس كورونا حوالي 300-500 نوكليوتيد (دون احتساب ذيل عديد الأدينين). النهاية 5' للمنطقة غير المترجمة 3' الخاصة بفيروسات بيتا (مباشرة بعد نهاية جين القفيصة النووية) عبارة عن حلقة جذعية منتبجة (BSL) ضرورية لتضاعف الفيروس طولها حوالي 70 نوكليوتيد، وقد تحتوي بعض فيروسات كورونا غاما على هذه البنية كذلك، ولكنها غير متواجدة في فيروسات ألفا. توجد بنية عقدة كاذبة طولها 54 نوكليوتيد مع الاتجاه بالنسبة للحلقة المنتبجة في فيروسات كورونا ألفا وبيتا، والتي قد تكون مطلوبة لتضاعف الفيروس. ترتبط حلقةٌ جذعية ثانية خاصة بالعقدة الكاذبة (PK-SL2) بتسلسلٍ في جذع الحلقة المنتبجة لتشكيل العقدة الكاذبة وهذا يعني أن الحلقة المنتبجة والعقدة الكاذبة متداخلتان في التسلسل ولا يمكنهما التطوي إلى هيئاتهما في آن واحد (صورة) وأنهما يعملان كمفتاح جزيئي. معدل تواجد الحلقة المنتبجة والعقدة الكاذبة بشكل مستقل أو تشكيلهما لهيئة مشتركة يكون بتوازن ديناميكي من أجل تنظيم بدء نسخ سلسلة الرنا الفيروسية سالبة الدلالة. فحين لا تشكل العقدة الكاذبة هيئة مشتركة مع الحلقة المنتبجة يُفترض أنها تترابط مع تسلسل النهاية 3' لتشكيل جذع يعمل كموقع لتجميع البروتينات اللابنيوية التي تعمل على تخليق السلسلة الجينومية السالبة، وعند الانتهاء من النسخ وانفصال تسلسل النهاية 3' تعود الحلقة الكاذبة للتشكل.
مع الاتجاه بالنسبة للعقدة الكاذبة تتواجد عقدة جذعية أخرى طويلة ومتفرعة غير ضرورية لتضاعف الفيروس تسمى المنطقة فائقة التغير (HVR) بسبب تغير تسلسلها الكبير بين الفيروسات. تحتوي هذه الحلقة على تسلسل ثماني محفوظ 5′-GGAAGAGC-3′ وظيفته غير واضحة لحد الآن.
البروتينات
يمكن تقسيم بروتينات فيروسات كورونا إلى قسمين: البروتينات البنيوية والبروتينات اللابنيوية، وتشمل البروتينات البنيوية:الغشاء (M) والغلاف (E) والشوكة (S) وفي بعض الفيروسات إستراز الراصة الدموية (HE) وتدخل جميعها في تركيب الفيروس. أما البروتينات اللابنيوية فهي 16 بروتينا تنتج من القص الذاتي لعديد البروتين 1ab الذي يُشفره جين الريبليكاز (ORF1ab) ولها أدوار مختلفة أهمها نسخ ومضاعفة الفيروس، بالإضافة إلى البروتينات البنيوية واللابنيوية، تملك العديد من فيروسات كورونا بروتينات ملحقة ذات وظائف متنوعة.
البروتينات البنيوية
بروتين الغشاء
بروتين الغشاء هو البروتين الرئيسي للغلاف الفيروسي والأكثر توافرا فيه من بين البروتينات البنيوية الأخرى، وهو بروتين غشائي من النوع الثالث طوله 218 إلى 263 حمض أميني ويشكل طبقة سمكها 7.8 نانومتر. ويملك نطاق نهاية أمنية قصيرخارجي وثلاث نطاقات عبر غشائية ونطاق نهاية كربوكسيلية داخلي يشكل مطرسا مثل الشبيكة يزيد من ثخانة ومتانة الغلاف. حسب الفيروس، يمكن أن تتم غلكزة النطاق الأميني الخارجي لهذا البروتين بربط الجليكانات في N أو في O. تلعب بروتينات الغشاء دورا مفتاحيا في تجميع الفيروس، فهي تغير بنيتها لتعديل انحناء الغشاء وتشكيل الغلاف الفيروسي، وتتآثر مع البروتينات البنيوية الأخرى لتنسيق تجميع الفيروس وتبرعمه.
تستخدم جميع فيروسات كورونا بروتين الشوكة للارتباط بالمستقبلات على سطح الخلية المضيفة، لكن أظهرت دراسات أن بروتين الغشاء الخاص بفيروس كورونا البشري NL63 يمكنه الارتباط بالجليكان البروتيني كبريتات الهيباران المتواجد على سطح الخلايا المضيفة، لكن يبقى بروتين الشوكة مطلوبا من أجل دخولٍ ناجح.
بروتين الغلاف
بروتين الغلاف هو بروتين بنيوي ثانوي يختلف كثيرا بين أنواع فيروسات كورونا. يوجد حوالي 20 بروتين غلاف فقط في فيروس كورونا، ويتراوح طوله بين 76 إلى 109 حمض أميني ووزنه الجزيئي بين 8.4 و12 كيلو دالتون. بروتين الغلاف هو الأصغر بين البروتينات البنيوية الأربعة وهو بروتين غشائي مدمج يملك نطاقين: نطاق عبر غشائي ونطاق نهاية كربوكسيلية خارجي، النطاق عبر الغشائي هو لولب ألفا مفرد يمكن أن يتجمع إلى خماسي قسيمات -كما هو الحال في سارس-كوف- ويشكل قنوات أيونية تسمى فيروبورينات تسمح للأيونات (بشكل أساسي أيونات الهيدروجين) بالمرور. وظيفة هذه القناة الأيونية مازالت قيد الدراسة، لكن قد يكون لها دور في عملية تحرير الفيروس من الخلايا. فضلا عن ذلك، تشارك بروتينات الغلاف أيضًا في تجميع الفيروس وتنقله داخل الخلية وتبرعمه. أظهرت الدراسات أن فيروس التهاب الكبد الفأري الذي يفتقر إلى بروتينات الغلاف لا يزال بإمكانه إصابة الخلايا. ولوحظ انخفاض في إمراضية فيروس السارس، أما ميرس-كوف الذي يفتقر إلى بروتين الغلاف فلم يكن قادرا على إصابة الخلايا. بالإضافة إلى ذلك، قد تثبط بروتينات الغلاف لبعض فيروسات كورونا الاستجابة المناعية للخلايا المضيفة.
بروتين الشوكة

بروتينات الشوكة هي السمة الأكثر تمييزا لفيروسات كورونا وهي المسؤولة عن مظهر السطح الذي يشبه الهالة أو التاج. يحتوي جسيم فيروس كورونا في المتوسط على 74 شوكة، ويبلغ طول كل شوكة حوالي 20 نانومتر وهي عبارة عن مثلوث من بروتين الشوكة. يتكون بروتين الشوكة بدوره من وحدتين فرعيتين S1 وS2. بروتين الشوكة الثلاثي المتجانس هو بروتين دمج غشاء من الصنف الأول يتوسط الارتباط بالمستقبلات واندماج الغشاء بين الفيروس والخلية المضيفة. تمثل الوحدة الفرعية S1 رأس الشوكة وتحتوي على نطاق الارتباط بالمستقبل، وتمثل الوحدة الفرعية S2 الجذع الذي يثبت الشوكة في الغلاف الفيروسي وتقوم عند تنشيطها بالبروتياز بدمج الغشائين. تظل الوحدتان الفرعيتان مرتبطتين بشكل غير تساهمي وهما مكشوفتان على سطح الفيروس حتى ترتبطان بغشاء الخلية المضيفة. ينقسم مركب الوحدات الفرعية إلى وحدات فرعية منفردة عندما يرتبط الفيروس بالخلية المضيفة ويندمج بها بفضل وظيفة البروتيازات مثل: عائلة الكاتبسينات، وبروتياز السيرين عبر الغشائي 2 (TMPRSS2) الخاصة بالخلية المضيفة.
نطاقات الوحدة S1 هي أهم مكونات الشوكة في إحداث الإصابة، وهي أيضا المكونات الأكثر تنوعا بين الفيروسات لأنها مسؤولة على الانتحاء الخلوي. تمتلك الوحدة الفرعية S1 نطاقين رئيسيين هما: نطاق النهاية الأمينية (S1-NTD) ونطاق النهاية الكربوكسيلية (S1-CTD)، وكلاهما يعمل كنطاق ارتباط بالمستقبل. تتعرف نطاقات النهاية الأمينية على السكريات الموجودة على سطح الخلية المضيفة وترتبط بها، بينما تتعرف نطاقات النهاية الكربوكسيلية على المستقبلات البروتينية المختلفة مثل الإنزيم المحول للأنجيوتنسين 2، أمينوببتيداز الألانين والببتيداز نازع الببتيديل 4 وترتبط بها.
فضلا عن النطاقات المستخدمة عادة، يمكن لجزيئات الغليكوزيل الموجودة على بروتين الشوكة أن ترتبط بالليكتينات الموجودة على سطح الخلية. على سبيل المثال: رغم أن سارس-كوف عادة ما يستخدم المسستقبل ACE2 إلا أن غليكوزيل بروتين اللشوكة يمكن أن يرتبط بـ CLEC4M وDC-SIGN وغيرها من الليكتينات الأخرى كمستقبلات بديلة.
قامت عدة دراسات بتحليل بنية بروتين الشوكة الخاص بفيروس كورونا الفأري ووجدت أن تطوي نطاق النهاية الأمينية لـS1 يشبه تطوي الجالكتين في الخلايا الحيوانية، ووفقا لذلك اقتُرحت فرضية أن أصل النطاق الأميني لبروتين الشوكة يرجع إلى خلية حيوانية مضيفة، أي أن أول فيروس كورونا حصل على الجين من الليكتين الخاص بالخلية المضيفة والذي يستطيع الارتباط بالجليكانات الموجودة على سطح الخلية واستخدمها كمستقبل لإصابة الخلايا، ولكن لاحقا تحور S1 الخاص بالعديد من فيروسات كورونا وتطور ليصبح قادرا على الارتباط بمستقبلات البروتين في الخلية المضيفة. على سبيل المثال: اتخذ النطاق الأميني لفيروس كورونا الفأري بنية جديدة واكتسب القدرة على الارتباط بـ CEACAM1 وهو ما زاد من قدرة الفيروس على الارتباط بخلايا الفئران بشكل كبير. ونظرا لعدم الحاجة إلى الارتباط بالسكريات فإنه فقد تدريجيا وظيفة الليكتين وزيادة على ذلك فقد وظيفة إستراز الراصة الدموية. في المقابل، فيروس كورونا البقري وفيروس كورونا البشري OC43 وغيرها مازالت تستخدم السكريات كمستقبلات، لذلك تحتفظ نطاقاتها الأمينية بوظيفة الليكتين. في عملية تطور S1 الخاص بفيروس كورونا، يواجه النطاق الكربوكسيلي ضغطا تطوريا أكبر، لذلك معدل التطور فيه أسرع من معدل التطور في النطاق الأميني. عندما يتحور النطاق الكربوكسيلي ويحاول الارتباط بمستقبلات جديدة، يمكن استخدام النطاق الأميني الذي يرتبط بالجليكوزيل كآلية تأمين لضمان إمكانية ارتباط الفيروس بالخلايا.
القص

يوجد موقعان في بروتين الشوكة يحدث فيهما القص من أجل أن يتمكن فيروس كورونا من إصابة الخلايا وهما: موقع S1/S2 عند حدود المنطقتين وموقع S2'. بعد القص في الموقع الأول تفقد المنطقتين S1 وS2 ترابطهما التساهمي، لكنهما تبقيان مرتبطتين ببعضهما عبر قوى بين جزيئية أخرى. عند الارتباط بالمستقبل تخضع بنية S2 لتغير هيئي يسمح بقص ثانٍ في موقع S2' يتحرر بواسطته ببتيد الدمج الذي يشرع في عملية دمج الغشائين الفيروسي والخلوي، وهو ما يسمح لجينوم الرنا الفيروسي بالدخول إلى السيتولازم.
تختلف فيروسات كورونا في مرحلة دورة حياة الفيروس التي يحدث فيها القص وخاصة في موقع S1/S2، إذ يُقص بروتين الشوكة في بعض فيروسات كورونا بواسطة بروتيازات داخل خلوية قبل أن يغادر الخلية. على سبيل المثال: يوجد تسلسل لدى فيروس كورونا الفأري A59،وميرس-كوف وسارس-كوف-2 بين S1 وS2 يمكن لبروتياز الفيورين أن يقوم بالقص فيه داخل الخلايا. في حين أن فيروسات أخرى تُقص بواسطة بروتيازات خارج خلوية مثل الإيلاستاز بعد الإخراج الخلوي. بعد الارتباط بالمستقبلات الخلوية تغير المنطقة S2 هيئتها بسرعة ثم يتم قصها في موقع S2' بواسطة بروتيازات أخرى على سطح الخلايا مثل بروتياز السيرين عبر الغشائي 2 بالنسبة لسارس-كوف-2، وبعد القص يمكن للفيروس أن يندمج مع غشاء الخلية ويدخل إلى السيتوبلازم.. الفيروسات التي لا تحتوي على مواقع قص للفيورين مثل سارس-كوف لا يتم قصها في S1/S1 وS2' سوى بواسطة بروتيازات على سطح الخلية أو في الجسيم الحال بعد إصابة الخلايا.
بروتين القفيصة النووية
توجد القفيصة النووية داخل الغلاف الفيروسي وتتكون من العديد من بروتينات القفيصة النووية المرتبطة بالرنا. يرتبط بروتين القفيصة النووية بجينوم الرنا مفرد السلسلة موجب الدلالة على هيئة خرز على خيط مستمرة، كما ترتبط بروتينات القفيصة النووية ببعضها البعض مشكلة خيوط قفيصة نووية عرضها من 10 إلى 15 نانومتر وطولها مئات النانومترات. بروتينات القفيصة هي بروتينات فوسفورية وزنها الجزيئي من 45 إلى 50 ألف كيلو دالتون وطولها في سارس-كوف 422 حمض أميني وفي سارس-كوف-2 419 حمض أميني. يتكون بروتين القفيصة من 5 نطاقات: نطاق أميني يرتبط بالرنا ونطاق كربوكسيلي يشكل مثنويات متجانسة، يفصلهما نطاق مركزي مضطرب جوهريا عالي الانحفاظ وغني بالسيرين، وعلى جانبيهما نطاقان آخران مضطربان جوهريا. بروتين القفيصة متعدد الوظائف وهو مولد كبير للمناعة حيث وُجدت أجسام مضادة له لدى المرضى المتعافين من مرضي السارس وكوفيد 19، وفضلا عن وظيفته الرئيسية في تعبئة الجينوم الفيروسي والمساهمة في عملية التجميع، يساهم بروتين القفيصة في عملية تبديل القالب لنسخ الرنا الجينومي الفرعي، ويدخل إلى النواة ويسبب توقف دورة الخلية في طور التركيب عبر تآثرات مع مركب الكيناز المعتمد على السيكلين كما أنه يثبط استجابات الإنترفيرون في كل من سارس-كوف، ميرس-كوف، وسارس-كوف-2 بطرق متنوعة.
إستراز الراصة الدموية
فضلا عن بروتينات الغشاء والغلاف والشوكة، يوجد بروتين آخر على غلاف فيروسات كورونا من جنس بيتا، الفرع A (مثل فيروس كورونا الفأري وفيروس كورونا البشري HKU1) هو إستراز الراصة الدموية (HE). هذا البروتين مثنوي ويتكون من وحدتين فرعيتين متماثلتين وطوله حوالي 400 حمض أميني، وهو بارز على سطح الفيروس مثل بروتين الشوكة لكنه أقصر منه وطوله حوالي 5 إلى 7 نانومتر، ووظيفته المساعدة في ارتباط الفيروس بالمستقبلات على الخلية المضيفة. بالإضافة إلى فيروسات كورونا، تحتوي فيروسات الإنفلونزا وفيروسات تورو على هذا البروتين كذلك، مما يشير إلى إمكانية حدوث تأشيب بين هذه الفيروسات. يتكون بروتين إستراز الراصة الدموية من نطاقين رئيسيين: نطاق ليكتين ونطاق إستراز. يمكن أن يرتبط كل من نطاق الليكتين وبروتين الشوكة بمستقبلات حمض السياليك على سطح الخلايا المضيفة، يساعد نطاق الإستراز في تحرير الفيروس من الخلية ومغادرته لها عبر قص أستيل من الوضعية 9 من جزيء حمض السياليك ( 9-O-Ac-Neu5Ac) النهائي. بعد انتقال فيروس كورونا البشري OC43 (الذي ينتمي إلى النوع فيروس كورونا بيتا 1) إلى مضيف بشري من فيروس كورونا البقري، فقد إستراز الراصة الدموية قدرة الارتباط بحمض السياليك تدريجيا، وأصبح بروتين الشوكة فقط هو الذي يربط بحمض السياليك، وتأثر نشاط نطاق الإستراز كذلك. خضع إستراز الراصة الدموية لفيروس كورونا البشري HKU1 لتطور تقاربي أيضا وحدثت له تغيرات مماثلة. قد تكون هذه التغيرات مرتبطة بتكيف الفيروس مع مستقبلات حمض السياليك على خلايا الجهاز التنفسي البشرية.
البروتينات اللابنيوية
يتكون إطار القراءة المفتوح ORF1ab من إطارين مفتوحين ORF1a وORF1b متصلين عبر انزياح إطار ريبوسومي مبرمج يسمح للريبوسوم بالاستمرار في الترجمة بعد كودون التوقف الموجود في نهاية ORF1a، ويسمى هذان الإطاران في بعض الأحيان بجين الريبليكاز. يشغل جين الريبليكاز المقبع 5' حوالي ثلثي الجينوم ويُترجم بترجمة معتمدة على القبعة. إلى عديدي البروتين pp1a وpp1ab الذين يخضعان لتحلل بروتيني تنتج منه عدة بروتينات لابنيوية ذات وظائف مختلفة في دورة حياة الفيروس، منها بروتيازات ومكونات مركب المضاعف الناسخ.
يحدث انزياح الإطار في تسلسل زلق تليه بنية رنا ثانوية على شكل عقدة كاذبة. وتم قياس نسبة فعالية انزياح الإطار بين 20-50% لدى فيروس كورونا الفأري، و45-70% لدى سارس-كوف-2، وهو ما يعني التعبير عن بروتين pp1a حوالي 1.5 إلى 2 مرة أكثر من البروتين pp1ab. يوجد في فيروسات كورونا 16 بروتينا لابنيويا، يحتوي عديد البروتين pp1a على البروتينات اللابنيوية من 1 حتى 11 ويحتوي عديد البروتين pp1ab على البروتينات اللابنيوية من 1 حتى 10 ومن 12 حتى 16. يقوم بالمعالجة التحللية (القص) بروتيازان: نطاق بروتياز مماثل للبابايين يتواجد في البروتين اللابنيوي 3 متعدد النطاقات ويقص البروتينات من 1 حتى 4، ثم يقوم البروتياز 3CL (ويُعرف بالبروتياز الرئيسي، وهو البروتين اللابنيوي 5) بباقي عمليات القص حتى النهاية الكربوكسيلية لعديد البروتين.
يسرد الجدول بالأسفل جميع الوظائف المعروفة لحد الآن للبروتينات اللابنيوية الخاصة بفيروساات كورونا:
| البروتين اللابنيوي | الوظيفة |
|---|---|
| البروتين اللابنيوي 1 | تفكيك الرنا الرسول الخلوي، تثبيط الترجمة الخاصة بالخلية المضيفة، تثبيط الإنترفيرون، غير متواجد لدى فيروسات كورونا غاما |
| البروتين اللابنيوي 2 | غير معروفة، يرتبط بالبروهيبتين |
| البروتين اللابنيوي 3 | بروتين متعدد النطاقات مع نطاق أو اثنين وظيفتهما ببروتياز مماثل للبابايين لقص البروتينات، مناهض للإنترفيرون، عدة أدوار أخرى. |
| البروتين اللابنيوي 4 | تكوين الحويصلات مزدوجة الغشاء |
| البروتين اللابنيوي 5 | بروتياز 3CL لمعالجة وقص عديد البروتين، تثبيط الإنترفيرون |
| البروتين اللابنيوي 6 | تكوين الحويصلات مزدوجة الغشاء |
| البروتين اللابنيوي 7 | عامل مرافق وعامل عملياتية لبوليمراز الرنا المعتمد على الرنا، يشكل مركبا مع nsp8 وnsp12. |
| البروتين اللابنيوي 8 | عامل مرافق وعامل عملياتية لبوليمراز الرنا المعتمد على الرنا، يشكل مركبا مع nsp7 وnsp12 |
| البروتين اللابنيوي 9 | يرتبط بالرنا مفرد السلسلة |
| البروتين اللابنيوي 10 | عامل مرافق للبروتينين nsp14 و nsp16. |
| البروتين اللابنيوي 11 | غير معروفة |
| البروتين اللابنيوي 12 | بوليمراز الرنا المعتمد على الرنا، وناقل النوكليوتيديل |
| البروتين اللابنيوي 13 | هيليكاز، وثلاثي فوسفاتاز الرنا. |
| البروتين اللابنيوي 14 | نوكلياز خارجي مصحح، تكوين قبعة الرنا، ناقل ميثيل-N7 غوانوزين. |
| البروتين اللابنيوي 15 | ريبونوكلياز داخلي، تجنب المناعة. |
| البروتين اللابنيوي 16 | ناقل ميثيل2'-O-ريبوز، تكوين قبعة الرنا. |
البروتينات الملحقة
فضلا عن الجينات المذكورة أعلاه، تحتوي معظم جينومات فيروسات كورونا على العديد من إطارات القراءة المفتوحة التي تشفر بروتينات ملحقة أو إضافية، وكلها تقع مع الاتجاه بالنسبة لجين الريبليكاز وهي موزعة بين أو داخل جينات البروتينات البنيوية الأربعة وبعضها له صلة بها. يمكن أن تكون إطارات القراءة المفتوحة الخاصة بالبرروتينات الملحقة متداخلة وتختلف أعدادها وأطوالها ومواضعها ووظائفها من فيروس إلى آخر، وهي في الغالب أصغر من البروتينات البنيوية وليست لازمة لتكاثر وتضاعف الفيروس وخاصة في الخلايا المختبرية. من وظائف البروتينات الملحقة تعديل الاستجابات المناعية والتهرب منها، لها دور في إجهاد الشبكة الإندوبلازمية والإستماتة والالتهام الذاتي، تعمل كفيروبورينات وتزيد من حدة الفيروس. على سبيل المثال تمنع البروتينات الملحقة استجابة الإنترفيرونات من النوع 1 وهي خط الدفاع الأول ضد الفيروسات بطرق واستراتيجيات متنوعة حيث يقوم ORF3b في سارس-كوف باستهداف MAVS ويقاطع ORF9b الخاص بسارس كوف-2 إضافة اليوبيكوتين إلى نيمو لتثبيط مسار RIG-I-MAVS ومنع تخليق الإنترفيرونات. يرتبط ORF6 الخاص بسارس-كوف بكاريوفيرين ألفا 2 لمنع نقل ستات 1 إلى النواة. ويثبط NS4a الخاص بميرس-كوف استجابات الإجهاد الفيروسية التي يتوسطها PKR.
دورة التضاعف
الدخول إلى الخلية

تستخدم معظم فيروسات كورونا بروتينات الشوكة للارتباط بالمستقبلات الموجودة على سطح الخلايا المضيفة، ولكن يمكن لبعض فيروسات كورونا أن ترتبط بالخلايا المضيفة ببروتينات أخرى مثل بروتين الغشاء الخاص بفيروس كورونا البشري NL63 وإستراز الراصة الدموية في الفيروسات الإمبيكوية. حيث يمكن لكلاهما أن يرتبط مع عديدات السكاريد على سطح الخلية للمساعدة في الإصابة. بعد ارتباط بروتين الشوكة بالمستقبل، تقوم بروتيازات المضيف (مثل الكاتبسين وبروتياز السيرين عبر الغشائي 2) بقصه وتنشيطه. تبعا لسلالة الفيروس، يمكن للفيروس النشط أن يدخل الخلية عن طريق الإدخال الخلوي أو يندمج مباشرة مع غشاء الخلية المضيفة ويدخل السيتوبلازم. يستخدم كل من: فيروس كورونا السنوري، 229E، فيروس كورونا الطيري، سارس-كوف، فيروس إلتهاب الكبد الفأري A59 وفيروس إلتهاب الكبد الفأري JHM الطريقة الثانية. فضلا عن البروتيازات، تؤثر بروتينات مضيفة أخرى على عملية إصابة الفيروس للخلية مثل: البروتين الغشائي المستحث بالإنترفيرون الذي يمنع الفيروسات من دخول الخلايا، والبروتين المحتوي على الفالوسين الذي يساعد الفيروسات الموجودة في الجسيمات الداخلية على دخول السيتوبلازم.
المستقبلات التي تستخدمها فيروسات كورونا للدخول إلى الخلية متنوعة وغير مرتبطة بجنس محدد فمثلا: ينتمي فيروس كورونا البشري NL63 إلى جنس فيروسات ألفا وسارس-كوف إلى فيروسات بيتا، لكن يستخدم كلاهما الإنزيم المحول للأنجيوتنسين 2 (ACE2) كمستقبل. أما فيروس التهاب المعدة والأمعاء الساري وفيروس الإسهال الوبائي الخنزيري وهما من نفس جنس فيروس NL63 (ألفا) يستخدمان مستقبل أمينوببتيداز الألانين. يستخدم فيروس كورونا الفأري وميرس-كوف اللذان ينتميان إلى فيروسات بيتا (مثل سارس-كوف) مستقبلي جزيء التصاق الخلية المرتبط بالمستضد السرطاني المضغي 1 والببتيداز نازع الببتيديل 4 على التوالي. علاوة على ذلك، تستخدم العديد من فيروسات كورونا جزيئات غليكوزيل معدلة موجودة في بروتينات على سطح الخلية كمستقبلات أو مستقبلات مساعدة. على سبيل المثال، يستخدم فيروس كورونا البشري OC43 وفيروس كورونا البقري 9-O-حمض السياليك كمستقبل.
ترجمة الجينوم

بعد دخول القفيصة إلى السيتوبلازم، تقص بروتيازات خلوية بروتينات القفيصة النووية لتحرير الجينوم الفيروسي. ولأن جينوم فيروس كورونا رنوي موجب الدلالة ويملك قبعة 5' وذيل عديد الأدينين مثل الرنا الرسول الخاص بحقيقيات النوى فإنه يُترجم مباشرة بواسطة الريبوسوم. أول ما تتم ترجمته هو جين الريبليكاز وفي سبيل التعبير عن كلا عديدي البروتين، يستخدم الفيروس تسلسلا زلقا 5'-UUUAAAC-3' وعقدة كاذبة لإحداث انزياح إطار ريبوسومي -1 للاستمرار في الترجمة وإنتاج عديد البروتين الكبير pp1ab. في معظم الحالات، يقوم الريبوسوم بفك بنية العقدة الكاذبة والاستمرار في الترجمة حتى يصادف كودون التوقف الخاص بـ ORF1a. لكن في بعض الأحيان وبمساعدة بروتينات فيروسية أو خلوية، تعيق العقدة الكاذبة الريبوسوم عن الاستمرار في الترجمة وتتسبب في توقفه مؤقتًا على التسلسل الزلق، وهو ما يؤدي إلى تغيُر إطار القراءة عبر العودة إلى الخلف بنيوكليوتيد واحد (انزياح إطار -1)، قبل أن يتمكن الريبوسوم من فك بنية العقدة الكاذبة والاستمرار في ترجمة ORF1b وإنتاج عديد البروتين الكامل pp1ab.
من غير المعروف بالضبط سبب استخدام الفيروسات لانزياح الإطار للتحكم في التعبير عن البروتينات، لكن افتُرض أن السبب إما من أجل التحكم الدقيق في نسبة بروتينات عديد البروتين pp1a وبروتينات عديد البروتين pp1b، أو لتأخير إنتاج بروتينات pp1b حتى تحضِّر بروتينات pp1a بيئة مناسبة لنسخ ومضاعفة الجينوم الفيروسي.
بعد ترجمة إطار القراءة ORF1a تقوم البروتينات اللابنيوية 3 و4 و6 بإحداث تغييرات في بنية الشبكة الإندوبلازمية وإنشاء عضيات التضاعف (ROs) التي يحدث فيها تضاعف الفيروس. تختلف فيروسات كورونا في نوع عضيات التضاعف التي تُحدثها وتشمل هذه العضيات: الأغشية الملففة (CM)، الحويصلات مزدوجة الغشاء (DMV)، الشبكة الإندوبلازمية المزمومة (zippered ER)، وتشير الدراسات إلى أن الحويصلات مزدوجة الغشاء هي مركز تضاعف الفيروس. تنشأ الحويصلات مزدوجة الغشاء من حويصلات مشتقة من الشبكة الإندوبلازمية، حيث تقوم بروتينات فيروسية بتسخير مسار تفكيك البروتين المرتبط بالشبكة الإندوبلازمية (ERAD) لإنشائها. كما يمكن أن تنشأ من استثارة مسار الالتهام الذاتي. يُعتقد أن القيام بعملية النسخ والتضاعف داخل الحويصلات مزدوجة الغشاء آلية لإخفاء الرنا الفيروسي ووسيلة لتجنب الاستجابات المضادة للفيروسات التي تُحدثها جزيئات الرنا مزدوجة السلسلة.
الناسخ المضاعف

في الحويصلات مزدوجة الغشاء، تتجمع العديد من البروتينات اللابنيوية (البروتين اللابنيوي 7 و8 و9 و10 و12 و13 و14 و16) التي نتجت من قص عديد البروتين pp1ab لتشكيل مركب التضاعف والنسخ لنسخ ومضاعفة جينوم الرنا الفيروسي. أهم بروتينات هذا المركب هو البروتين اللابنيوي 12 (بوليميراز الرنا، الذي يحفز تخليق الرنا، أما البروتينات الأخرى فلها وظائف مساعدة مختلفة مثل: البروتين اللابنيوي 14 (نوكلياز خارجي للرنا) الذي يوفر وظيفة تصحيح ويقوم بإزالة النيوكليوتيدات غير المتطابقة وتحسين دقة النسخ. تملك فيروسات الرنا عموما معدل طفرات عالٍ (10−5 إلى 10−3 طفرة لكل موقع لكل عملية نسخ) حين يتجاوز طول الجينوم 15 ألف نوكليوتيد وينتج عن ذلك حدوث كارثة خطأ بسبب تراكم العديد من الطفرات، لذلك جينومها له حد علوي. لكن معدل الطفرات في فيروس كورونا أقل بكثير من باقي فيروسات الرنا الأخرى بسبب وظيفة التصحيح الخاصة بالبروتين اللابنيوي 14 ويُقدر بـ (10−7 إلى 10 −6 طفرة لكل موقع لكل نسخ) وهو قريب من معدل الطفرات لفيروسات الدنا مفردة السلسلة، لذلك يمكن لجينوماتها بلوغ طول كبير يتجاوز 30 ألف نوكليوتيد. فضلا عن فيروسات كورونا، فيروسات الرنا الأخرى التي تملك وظائف التصحيح هي: فيروسات تورو والفيروسات الرونية وينتمي كلاهما إلى رتبة الفيروسات العشية وكلاهما يحتوي على جينومات أطول من جينومات فيروسات الرنا الأخرى. بالإضافة إلى هذه البروتينات اللابنيوية الفيروسية، يوجد العديد من بروتينات الخلية المضيفة في الغشاء الداخلي للحويصلات مزدوجة الغشاء بما في ذلك البروتينات المسؤولة عن النقل الحويصلي والبروتينات المرتبطة باليوبيكويتين والبروتينات المرتبطة بمسار الالتهام الذاتي والعديد من بروتينات بدء الترجمة التي تساعد في ترجمة الرنا. تبّين أن مركب التضاعف والنسخ يتم تجميعه بجوار الريبوسومات على غشاء الشبكة الإندوبلازمية لتحسين كفاءة ترجمة الرنا وتخليق البروتينات.
يتكون لب مركب التضاعف والنسخ في سارس-كوف-2 من بوليميراز الرنا المعتمد على الرنا (البروتين اللابنيوي 12، nsp12) وثلاث بروتينات لابنيوية: البروتين اللابنيوي 7 ونسختان من البروتين اللابنيوي 8. طول بوليميراز الرنا في سارس-كوف-2 932 حمض أميني ويتكون من أربع نطاقات رئيسية: نايران (NiRAN) ويملك وظيفة ناقل نوكليوتيديل، الواجهة، الأصابع والكف. ترتبط بالبوليميراز نسختان من البروتين اللابنيوي 8، وبسبب عدم تناظر البوليميراز فإن البروتين اللابنيوي 7 مطلوب من أجل أن يتوسط التآثرات بين البروتينين اللابنيويين 12 و8 على إحدى الجهتين. يرتبط هليكازان (البروتين اللابنيوي 13) بامتدادات لولبية للبروتينين 8 فوق لولب الرنا المزدوج (الصورة)، ويملك هذان الهليكازان تآثرات محدودة بين بعضهما البعض ويبدو أنهما يعملان على استقرار البنية الإجمالية لمركب التضاعف والنسخ. يعمل الهليكازان على تسهيل نسخ جينوم الرنا المليء بالبنى الثانوية والثالثية عبر تفكيك هذه البنى أما البروتينات اللابنيوية 9 و10 و14 و16 فلها أدوار في تنظيم تخليق القبعة 5' واستقرار بنى جينوم الرنا.
فضلا عن الهيئة الموحودة لبوليميراز الرنا، يمكن أن يشكل هذا الأخير مثنويا، يرتبط فيه كل بوليميراز بنسخة واحدة من البروتين 8 ويتثنيان عبر تآثرات بين البروتينين 7 الخاصين بهما ليتخذا هيئة توازي متضاد. افترض أن هذه الهيئة المثنوية تلعب دورا في نسخ الرنا الجينومي الفرعي وتبديل القالب.
الرنا الجينومي الفرعي وتبديل القالب
تملك الفيروسات العشية سمة مميزة لها عن باقي فيروسات الرنا وهي إنتاجها لرنا جينومي فرعي بأطوال مختلفة لتخليق البروتينات البنيوية والملحقة عبر آلية تبديل قالب معقدة. بعد تشكل مركب التضاعف والنسخ في الحويصلات مزدوجة الغشاء، يستخدم مركب التضاعف الرنا الفيروسي الموجب لإنتاج رنا سالب كامل الطول عبر نسخ مستمر، ثم يُستخدم هذا الرنا السالب كقالب لإنتاج رنا جينومي موجب كامل الطول. فضلا عن التضاعف الجينومي بالنسخ المستمر، يقوم مركب التضاعف بنسخ متقطع ينتج عنه رنا فرعي بأطوال مختلفة.
يتم إنشاء الرنا الفرعي عند إنشاء السلسلة السالبة عبر نموذج نسخ متقطع يفترض: وجود مواقع خاصة تسمى التسلسل المنظم للنسخ الجسمي (TRS-B) في بداية كل جين والتي تشكل تآثرات رنا-رنا مكملة بعيدة المدى مع التسلسل المنظم للنسخ الطليعي (TRS-L) المتواجد في حلقة الـSL3 في المنطقة غير المترجمة 5'. تُنظَّم هذه التآثرات بين التسلسل الجسمي والطليعي بواسطة مركبات رنا-بروتين يُفترض أنها تساهم في توقف مركب التضاعف والنسخ عند التسلسل الجسمي. بعد نسخ التسلسل الجسمي ينفصل مركب التضاعف والنسخ عنه ويعيد الارتباط بالتسلسل الطليعي مبدلا القالب ومتجاوزا جزءا كبيرا من الجينوم ويقوم بنسخ التسلسل الطليعي. بعد نسخ جزيئات الرنا الفرعية السالبة، تُستخدم هذه الأخيرة لنسخ رنا فرعي موجب، وعادة ما يُترجم من الرنا الجينومي الفرعي الموجب إطار القراءة المفتوح الأول عكس الاتجاه فقط، أما الرنا الجينومي السالب والرنا الجينومي الفرعي السالب فهما أقل تواترا في الخلايا المصابة وقد لا يشفران أي بروتينات فيروسية.
يُنظَّم تبديل القالب بواسطة بروتينات فيروسية وخلوية أهمها بروتين القفيصة النووية الذي يملك نشاط شابرون رنا يسهل عملية تبديل القالب وهو مطلوب ليكون النسخ فعالا، ومن بين وظائفه أنه يقوم بتوظيف الهليكاز الخلوي DDX1 الذي يسمح بتجاوز التوقف عند التسلسل الجسمي لنسخ رنا فرعي أطول أو رنا جينومي كامل. رغم أن معظم الرنا الفرعي (~90%) ينشأ عبر تبديل القالب بين التسلسلات الجسمية والتسلسل الطليعي إلا أن نسبة صغيرة منه (< 10%) يمكن أن تنشأ بطريقة مستقلة عن التسلسل الجسمي أو حتى مستقلة تماما عن التسلسلات المنظمة للنسخ، وهو ما يشير إلى حدوث تآثرات رنا-رنا غير قياسية تسببها بعض بُنى الرنا أو ارتباط عوامل فيروسية أو خلوية أثناء عمليات تبديل القالب، مثل تآثرات الرنوات النووية الصغيرة الخلوية (U1 وU2 وU4) مع مواقع محددة في إطاري القراءة ORF1a وORF1b.
التأشيب الجيني
لأن مركب التضاعف والنسخ بإمكانه تبديل القالب أثناء النسخ عند التسلسل المنظم للنسخ، فإنه يستطيع تبديل نسخ سلسلة فيروس بأخرى حين يتواجد أكثر من جينوم رنا فيروسي واحد في الخلية، وينتج عن ذلك تأشيب لجينات الرنا الفيروسي. تفاصيل الآلية مازلت غير معروفة، لكن أظهرت دراسة أن البروتين اللابنيوي 14 المسؤول عن التصحيح له دور كذلك في آلية التأشيب الجيني. التأشيب الجيني آلية مهمة في تنوع فيروسات كورونا، وتسمح بتطفير تسلسلات الفيروس لتوليد سلالات وأنواع جديدة وكذلك السماح للفيروسات بالانتقال عبر الأنواع. تملك عدة فيروسات كورونا سجلات على أحداث تأشيب جيني بينها، على سبيل المثال فيروس كورونا البشري OC43 المقسم إلى خمسة أنماط من A حتى E: النمط D هو ناتج تأشيب بين النمطين B وC، والنمط E هو ناتج تأشيب بين الأنماط B وC وD. تملك مستخلصة أمستردام-1 من فيروس كورونا البشري NL63 جزءا من تسلسل بروتين الشوكة الخاص بمستخلصة 496 وذلك بسبب تأشيب جيني، سارس-كوف (وهو من نمط بيتا) قد تكون له علاقة بالأنماط ألفا وغاما فقد وجدت دراسات علم تطور السلالات فيه مناطق تأشيب محتملة بين فيروسات من نمط ألفا وغاما وفيروسات أخرى من نمط بيتا، والعديد من فيروسات كورونا الخفاشية المرتبطة بالسارس عادة ما تقوم بتأشيبات جينية بين بعضها البعض. المصدر الرئيسي لنشوء فيروس السارس قد يكون حدوث تأشيبات بين عدة فيروسات كورونا خفاشية.
التجميع والتحرير

بعد نسخ الرنا الجينومي والفرعي تنتقل هذه الجزيئات إلى السيتوبلازم عبر مسامات موجودة في الحويصلات مزدوجة الغشاء، ويُترجم الرنا الفرعي لإنتاج البروتينات البنيوية والملحقة وتتموقع البروتينات البنيوية في الشبكة الإندوبلازمية وجهاز غولجي أين تتعرض لتعديلات ما بعد الترجمة، ثم تنتقل هذه البروتينات ومنها البروتين الشوكي في هيئة قبل الاندماج المثلوثية عبر حويصلات صغيرة إلى الحويصلات مفردة الغشاء (SMV) -وهي مكان تجميع الفيروس- وتندمج بها. ثم يتجمع بروتين الشوكة والغشاء والغلاف على جزءٍ من غشاء الحويصلة مفردة الغشاء وتتسبب تآثرات بينه وبين بروتينات الغلاف والغشاء في انحنائه. وبوجود القفيصة الفيروسية وتآثرات بين بروتينات الغشاء والقفيصة يحدث تبرعم الفيروس إلى داخل الحويصلة مفردة الغشاء التي تحتوي في النهاية على العديد من الفيروسات ثم تغادر هذه الأخيرة الخلية عبر الإخراج الخلوي. بعدها يمكن للفيروسات الجديدة المحررة إصابة خلايا أخرى. عملية تجميع فيروس كورونا يمكن أن تساهم فيها بروتينات من الخلية المضيفة مثل الأنيبيبات الدقيقة، خيوط الأكتين والفيمنتينات، وتختلف هذه البروتينات حسب الفيروس.
يقوم بروتين القفيصة النووية بتغليف الرنا الجينومي لتكوين القفيصة النووية وتكديسها من أجل تعبئتها في الفيروس من خلال التآثر مع إشارة التغليف التي يختلف موقع تواجدها حسب نوع الفيروس ولا توجد سوى في الرنا الجينومي الفيروسي الكامل وهي الطريقة التي يتجنب بها الفيروس تغليف رنا جينومي فرعي أو رنا يخص الخلية المضيفة، ويمكن لكلا بروتيني الغشاء والقفيصة التعرف على هذه الإشارة والتآثر معها. آلية تكوين القفيصة وتجميع الفيروس غير واضحة تماما، لكن تشير دراسات على سارس-كوف-2 إلى أن بروتين القفيصة عند ارتباطه بالرنا الجينومي يخضعان لانفصال طور سائل-سائل ويشكلان تكثفات أو تكدسات. يتكون بروتين القفيصة من نطاقين منحفظين: نطاق أميني يرتبط بالرنا يُعتقد أنه يقوم بتآثرات مع إشارة التغليف ونطاق كربوكسيلي يشكل مثنوي متجانس يُفترض أن له دور في تجميع البروتين النووي الريبوزي (تكوين القفيصة)، وتتواجد وسط وعلى جانبي هذين النطاقين نطاقات مضطربة جوهريا (IDRs) (الصورة)، النطاق المركزي عالي الانحفاظ وغني بالسيرين وفسفرته تلعب دورا في سلوك الهلام أو السائل لتكثفات بروتين القفيصة والرنا. النطاقان الجانبيان أقل انحفاظا والنطاق الكربوكسيلي منهما الخاص بسارس-كوف يرتبط بالبروتين الغشائي وهو ما يوحي بدور مهم لهذا النطاق في تعبئة وتجميع الفيروس.
أشارت دراسات سابقة إلى أن البروتينات النووية الريبوزية الفيروسية (vRNP) خيطية لولبية الشكل لكن أظهرت دراسات بالتصوير المقطعي البردي الإلكتروني (cryo-ET) على سارس-كوف-2 بُنى حبيبية الشكل، حيث يحتوي كل فيروس على 35-40 مركب بروتيني نووي ريبوزي يتخذ كل واحد منها بنية أسطوانية صدفية قطرها 15 نانومتر. وداخل الفيروس، تتجه هذه المركبات القريبة من الغشاء الفيروسي باتجاه الغشاء، الأمر الذي يوحي بأنها تتآثر بشكل خاص مع البروتين الغشائي. وداخل مركب البروتين النووي الريبوزي، تقترح نمذجة تجريبية مستندة على بُنى النطاقين الأميني والكربوكسيلي أنه يتكون من حوالي 800 نوكليوتيد مغلفة بحوالي 12 نسخة من بروتين القفيصة، يمكن لمركبات البروتين النووي الريبوزي المفردة أن تشكل تكدسات خطية وهذا ما يفسر التضارب في الشكل الملاحظ لها ويوحي بأنها يمكن أن تتخذ أشكالا منحفظة متنوعة.
الانتقال
الأفراد المصابون قادرون على نشر الفيروسات في البيئة. تآثر بروتين الشوكة مع مستقبل خلوي مكمل له أمر أساسي في تحديد انتحاء الأنسجة، شدة العدوى، ومجال الأنواع التي يصيبها الفيروس. تستهدف فيروسات كورونا بشكل أساسي الخلايا الظهارية، وتنتقل من مضيف إلى مضيف آخر حسب نوع الفيروس إما عن طريق الرذاذ التنفسي أو المواد المحتوية على الفيروس، أو الطريق الفموي الشرجي.
تصيب فيروسات كورونا البشرية الخلايا الظهارية في الجهاز التنفسي، بينما تصيب فيروسات كورونا الحيوانية عموما الخلايا الظهارية في الجهاز الهضمي. على سبيل المثال، يصيب فيروس كورونا السارس الخلايا الظهارية للرئة البشرية عن طريق الهباء الجوي بالارتباط بمستقبل الإنزيم المحول للأنجيوتنسين. ويصيب فيروس التهاب المعدة والأمعاء الساري الخلايا الظهارية للخنازير في الجهاز الهضمي عبر الطريق الفموي الشرجي بالارتباط بمستقبل أمينوببتيداز الألانين.
استجابة الخلية المضيفة
الالتهام الذاتي
بعد الإصابة بفيروس كورونا يمكن للبروتينات اللابنيوية أن تستثير الالتهام الذاتي، إذ ترتبط عملية تكوين الحويصلات مزدوجة الغشاء (DMV، موقع تضاعف الرنا الفيروسي) في الشبكة الإندوبلازمية ارتباطا وثيقا باستجابة الالتهام الذاتي، وتتشكل بعد إعادة تنظيم أو تغيير بنية غشاء الشبكة الإندوبلازمية. مع ذلك، توجد نتائج تجريبية تفيد بأن فيروسات كورونا يمكنها أن تصيب بنجاح الخلايا التي تعوز بروتينات الالتهام الذاتي المهمة مثل ATG5. لذلك يمكن لبعض فيروسات كورونا (مثل: فيروس التهاب الكبد الفأري، وفيروس التهاب الشعب الهوائية المعدي، وفيروس كورونا البشري NL63) أن تحفز تكوين الحويصلات مزدوجة الغشاء من خلال مسار آخر (مسار ERAD) وهو مسار مستقل عن مسار الالتهام الذاتي، لكنه يستعين ببعض البروتينات المستخدمة في مسار الالتهام الذاتي مثل MAP1LC3B. في استجابة الالتهام الذاتي، يقوم BECN1 بتحفيز اندماج الجسيمات البلعمية الذاتية واليحلولات لتفكيك المواد الموجودة فيها، ويمكن لفيروسات كورونا أن تمنع عمل BECN1 بعدة طرق لتجنب اندماج الحويصلات مزدوجة الغشاء مع اليحلولات.
استجابة إجهاد الشبكية الإندوبلازمية
تؤدي عملية تضاعف فيرووس كورونا وتجميعه إلى تغيّر ملحوظ في بنية الشبكة الإندوبلازمية، حيث تترجَم جميع بروتينات الفيروس بواسطة الريبوسومات في الشبكة الإندوبلازمية ويخضع معظمها إلى تعديلات بعد ترجمة متنوعة منها: الغلكزة، إضافة البالميتيك إضافة اليوبيكويتين، إضافة الـADP ريبوز، إضافة بروتين سومو، كما أن تطوي وبلمرة هذه البروتينات يعتمد على مساعدة الشابرونات الموجودة داخل الشبكة الإندوبلازمية وهو ما يزيد العبء على هذه الأخيرة. فضلا عن ذلك، تؤدي الإصابة بالفيروس إلى استثارة مسار الالتهام الذاتي ومسارات أخرى تُنتِج الكثير من الحويصلات التي تستهلك مكونات غشاء الشبكة الإندوبلازمية وهو ما يُحدث ضغطا على الشبكة. تؤدي جميع هذه العوامل إلى إثارة استجابة خلوية تسمى استجابة إجهاد الشبكية الإندوبلازمية. تُحفز استجابة إجهاد الشبكة عدة مسارات خلوية تقوم بتقليل ترجمة البروتينات وتعمل كآلية لمحاربة الإصابة بالفيروس، ففي بعض فيروسات كورونا يُنشِّط إجهاد الشبكة كينازات عامل البدء 2 مثل: EIF2AK3 وPKR في المرحلة الأولى من الإصابة والتي تقوم بفسفرة عامل البدء eIF2 لمنع الترجمة وتحفير الاستماتة وتخليق السيتوكينات. على سبيل المثال: يمكن تحفيز التعبير عن الإنترفيرون من النوع 1 بعد أن تنشِّط الإصابة بفيروس التهاب المعدة والأمعاء الخنزيري مسار EIF2AK3. يمكن لإجهاد الشبكة كذلك أن ينشط مسار IRE1-XBP1 ومسار ATF6، والعديد من المسارات المختلفة التي تؤدي إلى تخليق البروتينات المضادة للإجهاد والسيتوكينات.
الاستماتة والسيتوكينات
يمكن لفيروسات كورونا أن تُحدِث الاستماتة من خلال عدة آليات، إذ يمكن أن يسبب سارس-كوف استماتة خلايا الرئة والطحال والغدة الزعترية وأنسجة أخرى من خلال مسار الكاسباز، ويمكن أن يسبب ميرس-كوف الاستماتة لأنسجة الجهاز التنفسي والخلايا التائية بطرق مختلفة، كما يمكن أن يتسبب فيروس كورونا البشري OC43 في استماتة الخلايا العصبية بمسار مستقل على الكاسباز لكن يستخدم بروتين Bax . ويُحدث سارس-كوف-2 الاستماتة كذلك. يمكن أن تتسبب الإصابة بفيروس كورونا أيضا بشكل غير مباشر في استماتة الخلايا المصابة عن طريق تحفيز استجابة إجهاد الشبكة الإندوبلازمية أو تحفيز مسار MAPK/ERK. فضلا عن أنسجة الانتحاء الرئيسية، تسبب الفيروسات المذكورة سابقا موت العديد من أنواع الخلايا المناعية. إذ يمكن لفيروس كورونا البشري 229E أن يقتل الخلايا المتغصنة (ولكن ليس من خلال الاستماتة)، وبالتالي يمكن أن يثبط الاستجابة المناعية للمضيف. هناك العديد من الطرق للتسبب في الاستماتة بما في ذلك مسارات استجابة إجهاد الشبكة الإندوبلازمية المذكورة آنفا، والعديد من مسارات MAPK التي يمكن أن تسببها عدوى فيروس كورونا. على سبيل المثال، بعد تنشيط كينازات p38، يمكن فسفرة EIF4E لتثبيط الترجمة وتحفيز الاستماتة، تحفز كينازات JNK الاستماتة كذلك وقد يكون لها وظائف أخرى مضادة للفيروسات. مع ذلك، يوجد مسار MAPK آخر يُحدثه فيروس كورونا هو مسار ERK له نتائج عكس نتائج مسار كينازات p38، إذ يقوم بفسفرة eIF4EBP1 وتحرير eIF4E المرتبط به ليقوم بدوره في بدء عملية الترجمة.
يمكن للإصابة بفيروس كورونا أن تتسبب في تخليق مختلف السيتوكينات عبر مسارات مختلفة مما يؤدي إلى استجابة التهابية، وهي الاستجابة المناعية الفطرية للخلايا المضيفة. فضلا عن المسارات المذكورة أعلاه التي تحفز التعبير عن السيتوكينات نتيجة إجهاد الشبكة الإندوبلازمية، يمكن لفيروسات كورونا أيضا أن تنشط العديد من مسارات MAPK في الخلايا مثل: مثل p38 وERK وJNK ويمكن لهذه المسارات الثلاثة تحفيز التعبير عن مجموعة متنوعة من السيتوكينات. تم الإبلاغ عن سيتوكينات مثل: CCL2 وإنترلوكين 8 وإنترلوكين 6 وTNF-α وCOX-2 في مختلف فيروسات كورونا.
المناعة التكيفية
بعد إصابة المضيف، يمكنه تطوير مناعة تكيفية ضد فيروس كورونا عبر تنشيط خلايا تائية وخلايا بائية محددة. يمكن أن تستمر مناعة الأشخاص الأصحاء ضد فيروسات كورونا الأربعة (229E وOC43 وNL63 وHKU1) التي تسبب الزكام لنحو نصف عام إلى عام فقط، وبعد ذلك يمكن أن يصابوا بنفس الفيروس مرة أخرى. يتم إنتاج الأجسام المضادة ضد بروتين الشوكة الخاص بسارس-كوف في غضون ثلاثة أسابيع، وقد يكون الوقت اللازم لإنتاجها في سارس-كوف-2 أقصر.عيار الأجسام المضادة لمرضى السارس سوف يتغير بمرور الوقت، تتبعت بعض الدراسات المرضى في حدث السارس ووجدت أن استجابات الأجسام المضادة وخلايا البائية الذاكرة في المرضى انخفضت بشكل ملحوظ بعد عدة سنوات. في المقابل، يمكن أن يحافظ الجسم على خلايا التائية الذاكرة لفترة أطول. مع ذلك، قد تختلف مدة المناعة من شخص لآخر حيث وُجد أنه لا يزال بالإمكان اكتشاف الأجسام المضادة لدى بعض المرضى بعد إصابتهم بالسارس منذ أكثر من عشر سنوات. تختلف مدة بقاء الأجسام المضادة التي تنتجها الإصابة بفيروس ميرس-كوف تبعا لشدة الأعراض، فعيار الأجسام المضادة بعد سنتين من الإصابة أكبر بكثير لدى مرضى الحالات الحادة منه لدى مرضى الحالات المتوسطة.
يدوم مفعول التطعيم ضد فيروس التهاب الشعب الهوائية المعدي في الدجاج حوالي 9 أسابيع فقط وكان فعالا ضد بعض السلالات فقط . يشبه فيروس كورونا التنفسي الخنزيري (PRCV) إلى حد كبير فيروس التهاب المعدة والأمعاء الساري الخنزيري، ويمكن للخنازير المصابة به اكتساب مناعة ضده فقط وليس لكليهما، لكن المناعة المخاطية ضده (الغلوبولين المناعي أ) تضعف بمرور الوقت، الأبقار المصابة بفيروس كورونا البقري تضعف مناعتها بمرور الوقت كذلك. تؤدي الاستجابة المناعية الناتجة عن الإصابة بسلالة JHM من فيروس التهاب الكبد الفأري إلى إزالة الميالين من الخلايا العصبية في الدماغ مع القضاء على الفيروس.
آليات مقاومة الفيروس للمناعة

تمتلك فيروسات كورونا مجموعة متنوعة من الآليات لمجابهة الاستجابة المناعية للخلايا المضيفة. فضلا عن وظائفها البنيوية أو التحفيزية، تملك العديد من بروتينات فيروس كورونا وظائف تهرب ومقاومة للاستجابة المناعية. ومقارنة بفيروسات الرنا الأخرى لا يسبب فيروس كورونا قدرا كبيرا من التعبير عن الإنترفيرونات من النوع الأول، مما يوحي بأن لديه آليات لمنع التعبير عنها. يملك البروتين اللابنيوي 3 (بروتياز PLpro) الخاص بفيروسات: سارس-كوف، ميرس-كوف، IBV، كورونا الفأري، NL63 البشري -زيادة على وظيفة القص الذاتي- وظيفة إزالة اليوبيكويتين من بروتينات المضيف، ويملك نفس البروتين في سارس-كوف وميرس-كوف وظيفة إضافية هي إزالة وسم ISG15 من بروتينات المضيف. ولكون إضافة كلٍّ من اليوبيكويتين وISG15 تلعب دورا مهما في مسارات التأشير الخاصة بالاستجابة المناعية الخلوية، فإن إزالة هذه الوسوم تثبط الاستجابة المناعية وتخفض من التعبير عن الإنترفيرونات وهي الطريقة التي يقاوم بها البروتين اللابنيوي 3 الاستجابة المناعية، فضلا عن آليات المقاومة بالقص هذه يملك نفس البروتين في سارس-كوف آلية لتثبيط تخليق الإنترفيرون 1 مستقلة عن القص بالبروتياز. يمكن لكل من البروتين اللابنيوي 3 والبروتين اللابنيوي1 وORF3b وبروتين القفيصة النووية في سارس-كوف أن يثبطوا تنشيط مسار RIG-I لمنع الرنا الفيروسي مزدوج السلسلة من إثارة استجابة مناعية، ويمكن لـORF6 أن يثبط تنشيط مسار JAK/STAT بحيث تثبط كلا الآليتين تخليق الإنترفيرون 1. يمنع البروتين اللابنيوي 1 الكثير من جزيئات الرنا الرسول الخاصة بالمضيف -ومن بينها الرنا الرسول الخاص بالإنترفيرونات- من الترجمة ويسبب تفككها، أما الرنا الرسول الخاص بالفيروس في حد ذاته فهو لا يتأثر بوظيفة البروتين اللابنيوي 1 بفضل تسلسل خاص في المنطقة غير المترجمة 5'.
فضلا عن تنشيط RIG-I، يمكن للرنا الفيروسي مزدوج السلسلة أن ينشط 2'-5' مخلقة قليل الأدينيلات لتخليق 2'-5' قليل الأدينيلات وبعدها تنشيط الريبونوكلياز L، وفي سبيل مقاومة هذه الاستجابة يقوم البروتين اللابنيوي 4B (ns4b) في ميرس-كوف والبروتين اللابنيوي 2 في فيروس التهاب الكبد الفأري بتفكيك 2'-5' قليل الأدينيلات لمنعه من تنشيط الريبونوكلياز L وبالتالي منع الرنا الفيروسي من التفكك. يمكن لبروتين الغلاف في سارس-كوف أن يثبط مسار ERN1 الذي يُحفز بواسطة استجابة إجهاد الشبكة الإندوبلازمية وبالتالي يمنع الاستماتة وتخليق السيتوكينات. وربما هذه الآلية قد تكون مرتبطة بوظيفة بروتينات الغلاف في تشكيل فيروبورينات (قنوات آيونية). يقوم بروتين القفيصة النووية بتكوين تكثفات مع الرنا ويستخدم بروتين لب حبيبة إجهاد G3BP1 وبالتالي يحرم الخلية من استخدامه في تكوين حبيبات الإجهاد ويمنع الاستجابات المناعية الفطرية المعتمدة على G3BP1.
التصنيف
فيروسات كورونا هي عائلة فرعية تنتمي إلى عائلة الفيروسات التاجية، التي تنتمي إلى الرتبة الفرعية «كورنيدوفرينيات» (Cornidovirineae) التي تنتمي إلى رتبة الفيروسات العشية. تنقسم عائلة الفيروسات التاجية إلى ثلاث فروع: الكورونافيرينات المستقيمة (Orthocoronavirinae) والتي تعرف بالاسم الشهير فيروسات كورونا، الليتوفيرينات، والعائلة الفرعية الجديدة البيتوفيرينات (Pitovirinae). تنقسم عائلة فيروسات كورونا الفرعية إلى أربع أجناس (فيروس كورونا ألفا، فيروس كورونا بيتا، فيروس كورونا غاما، فيروس كورونا دلتا) وتصيب أجناس ألفا وبيتا الثدييات فقط، أما غاما ودلتا فيصيب معظمها الطيور بشكل رئيسي مع أنواع قليلة تصيب الثدييات. ينتمي فيروسا سارس-كوف وسارس-كوف-2 إلى نوع الفيروسات المرتبطة بالسارس التي تنتمي إلى الجنس الفرعي الفيروسات الساربيكوية التي تنتمي إلى جنس فيروسات بيتا. يعتمد تصنيف فيروسات كورونا على سبع تسلسلات محفوظة في جين ORF1ab فإذا كانت تسلسلات الأحماض الأمينية الناتجة من هذه التسلسلات السبع متماثلة بين سلالتين من الفيروس بنسبة تزيد عن 90% تعتبر السلالتين من نفس النوع.
فيروس كورونا ألفا
النوع النموذجي لفيروس كورونا ألفا هو فيروس كورونا ألفا 1 (ألفا-كوف-1)، الذي ينقسم إلى 14 جنسا فرعيا منها: فيروس كورونا ألفا 1 (فيروس كورونا الكلبي، فيروس كورونا القططي، فيروس التهاب المعدة والأمعاء الساري الخنزيري)، فيروس كورونا ألفا 2 (فيروس كورونا ابن مقرض، فيروس كورونا المنك)، فيروس كورونا الألبكة، فيروس كورونا البشري 229E، فيروس كورونا البشري NL63، فيروس كورونا خفاش الحذوة الكبير HuB-2013 (BtRf-AlphaCoV/HuB2013)، فيروس كورونا الخفاش محني الجناح نوع 1 (Bat-CoV MOP1)، فيروس كورونا الخفاش محني الجناح HKU8 (Bat-CoV HKU8)، فيروس كورونا خفاش مايوتيس ريكيتي Sax-2011 (BtMr-SAX2011)، فيروس كورنا الخفاشي CDPHE15 (BtCoV CDPHE15)، فيروس كورنا الخفاشي 3398 (PK-BatCoV 3398)، فيروس كورونا خفاش نيكتالوس فيلتينوس SC-2013 (BtNv-SC2013)، فيروس كورونا زبابات T14 (Sa-CoV T14)، فيروس كورونا خفاش إلف الظلام 512 (Bat-CoV 512)، فيروس الإسهال الوبائي الخنزيري (PEDV)، فيروسات خفاش الأقحوان مثل فيروس كورونا خفاش حدوة الفرس الصيني HKU2 (Bat-CoV HKU2)، فيروس كورونا متلازمة الإسهال الحاد الخنزيري (SADS-CoV)، فيروس كورونا زبابة ونتشنغ (WESV) وفيروس كورونا الجرذ البني لوتشنغ (LRNV).
فيما يلي الشجرة التطورية لكل مجموعة رئيسية من فيروس كورونا ألفا (راجع أمبر بابينو وزملائها، تشيتشيانغ وو وزملائه):
| فيروس كورونا ألفا |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
فيروس كورونا بيتا

النوع النموذجي لجنس فيروس كورونا بيتا هو فيروس كورونا الفأري (م-كوف) الذي ينقسم إلى 5 أجناس فرعية منها: فيروس كورونا بيتا 1 (فيروس كورونا البشري OC43، فيروس كورونا البقري، فيروس كورونا الخيلي، فيروس التهاب الدماغ والنخاع الشوكي الراصي الخنزيري)، فيروس كورونا الأرنبي HKU14 (RbCoV HKU14)، فيروس كورونا الجرذ الصيني HKU24 (ChRCoV HKU24)، فيروس كورونا البشري HKU1، فيروس كورونا الفأري، فيروس كورونا ميودس 2JL14 (MrufCoV 2JL14)، فيروس كورونا خفاش أيدولون C704 (Ei-BatCoV C704)، فيروس كورونا خفاش بيبيسترالوس HKU5 (Bat-CoV HKU5)، فيروس كورونا خفاش راوستوس HKU9 (HKU9-1)، فيروس كورونا خفاش راوستوس GCCDC1 (Ro-BatCoV GCCDC1)، فيروس كورونا خفاش تيلونيكترايس HKU4 (Bat-CoV HKU4)، فيروس كورونا المرتبط بمتلازمة الشرق الأوسط التنفسية (ميرس-كوف)، فيروس كورونا القنفذ البري (EriCoV)، فيروس كورونا خفاش زاريا (ZBCoV)، فيروس كورونا خفاش برات مستدير الورق جيجيانغ2013 (BtHp- BetaCoV/ZJ2013)، وفيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة (سارس-كوف، سارس-كوف-2 والسلالات المرتبطة) والعديد من الفيروسات الأخرى.
فيما يلي الشجرة التطورية للمجموعات الرئيسية من فيروس كورونا بيتا (راجع أمبر بابينو وزملائها ، تشيتشيانغ وو وزملائه):
| فيروس كورونا بيتا |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
فيروس كورونا غاما
النوع النموذجي لجنس فيروس كورونا غاما هو فيروس كورونا الطيري (AvCoV) وينقسم إلى ثلاثة أجناس فرعية منها: فيروس كورونا الحوتي، فيروس كورونا الطيري، فيروس كورونا الطيري 9203 (AvCoV 9203)، فيروس كورونا الإوزي CB17 (BcanCoV CB17)، فيروس كورونا البطي 2714 (DuCoV 2714)، وفيروسات أخرى.
فيما يلي الشجرة التطورية للمجموعات الرئيسية من فيروس كورونا غاما (راجع أمبر بابينو وزملائها ، ميشال ويلي وزملائها):
|
فيروس كورونا دلتا
النوع النموذجي لجنس فيروس كورونا دلتا هو فيروس كورونا البلبلي HKU11 (Bulbul-CoV HKU11) وينقسم إلى ثلاث أجناس فرعية منها: فيروس كورونا ودجون HKU20 (WiCoV HKU20)، فيروس كورونا البلبلي HKU11، فيروس كورونا الخنزيري HKU15 (PorCoV HKU15)، فيروس كورونا دجاج الماء الشائع HKU21 (CMCoV HKU21)، فيروس كورونا مونيا HKU13 (MuCoV HKU13)، فيروس كورونا أبيض العين HKU16 (WeCoV HKU16) فيروس كورونا العقعق روبن HKU18 (MRCoV HKU18)، فيروس كورونا بلشون الليل HKU19 (NHCoV HKU19)، فيروس كورونا الصقري HKU27 (FalCoV UAE-HKU27)، فيروس كورونا الحبارى الإفريقي HKU28 (HouCoV UAE-HKU28)، فيروس كورونا الحمام HKU29 (PiCoV UAE-HKU29)، فيروس كورونا السمان HKU30 (QuaCoV-HKU30) وفيروسات أخرى.
فيما يلي الشجرة التطورية للمجموعات الرئيسية من فيروس كورونا دلتا (راجع أمبر بابينو وزملائها، وميشال ويلي وزملائها):
|
التطور

تستخدم بعض الدراسات بوليميراز الرنا المعتمد على الرنا كساعة جزيئية وتقدر أن أحدث سلف مشترك لجميع فيروسات كورونا برز إلى الوجود بين 8000-10000 سنة قبل الميلاد، وأن أحدث سلف مشترك لفيروسات كورونا ألفا يعود إلى 2400 سنة قبل الميلاد ولفيروسات بيتا إلى سنة 3300 سنة قبل الميلاد ولفيروسات غاما إلى 2800 سنة قبل الميلاد ولفيروسات دلتا إلى 3000 سنة قبل الميلاد. لكن بعض العلماء يعتقدون أن هذه الدراسات تبالغ في تقدير معدل التطفر لفيروسات كورونا وتتجاهل تأثير اصطفاء التنقية ويقترحون نموذجا يزعم أن أحدث سلف مشترك لفيروسات كورونا ظهر منذ 55 مليون سنة وخضع إلى تطور مشترك طويل الأمد في الخفافيش والطيور. الخفافيش والطيور مستودع طبيعي مثالي لجينات فيروس كورونا لكونها فقاريات طائرة من ذوات الدم الحار (مع كون الخفافيش مستودعا لفيروسات ألفا وبيتا، والطيور مستودعا لفيروسات غاما ودلتا). مكّن العدد الكبير لأنواع الخفافيش والطيور التي تستضيف الفيروسات والنطاق العالمي لانتشارها من تطور كثيف وانتشار كبير لفيروسات كورونا.
يرجع أصل العديد من فيروسات كورونا البشرية إلى الخفافيش. اشترك فيروس كورونا البشري NL63 في سلف مشترك مع فيروس كورونا الخفاشي(ARCoV.2) بين 1190-1449 م. وتشارك فيروس كورونا البشري 229E مع فيروس كورنا الخفاشي (GhanaGrp1 Bt CoV) في سلف مشترك بين 1686-1800 م. حديثا، تشعب فيروس كورونا الألبكة عن فيروس 229E في وقت ما قبل عام 1960. انتقل ميرس-كوف إلى البشر من الخفافيش عبر الجمال كمضيف وسيط. ومع أن ميرس-كوف له صلة بالعديد من فيروسات كورونا، يبدو أنه تشعب عن هذه الأخيرة منذ عدة قرون. تشعب سارس-كوف عن أقرب فيروسات كورنا الخفاشية صلة به سنة 1986. أصاب أسلاف سارس-كوف أول مرة خفاش الأنف الورقي من عائلة الهيبوسيدرديات ثم انتشرت إلى خفاش حدوة الفرس في عائلة خفاشيات حدوة الفرس، ثم إلى زباد النخيل الآسيوي وأخيرا إلى البشر.
خلافا لغيره من فيروسات كورونا بيتا، يرجع أصل فيروس كورونا البقري الذي ينحدر من النوع فيروس كورونا بيتا 1 والجنس الفرعي فيروس أمبيكو إلى القوارض وليس إلى الخفافيش. في عقد 1790 تشعب فيروس كورونا الخيلي عن فيروس كورونا البقري بعد انتقال عبر الأنواع. ولاحقا في العقد 1890 تشعب فيروس كورونا البشري OC43 عن فيروس كورونا البقري بعد حادثة انتقال أخرى عبر الأنواع. افتُرض أن جائحة الإنفلونزا 1890 قد يكون سببها حادثة انتقال عبر الأنواع، وليس فيروس الإنفلونزا بسبب توقيت حدوثها، الأعراض العصبية والمسبب غير المعروف للجائحة. فضلا عن التسبب في عداوى تنفسية، اشتُبه أن فيروس كورونا البشري OC43 لعب دورا في الأمراض العصبية. في عقد 1950 بدا فيروس OC43 في التشعب إلى أنماطه الجينية الحالية. في علم تطور السلالات، فيروس التهاب الكبد الفأري الذي يصيب كبد الفأر والجهاز العصبي المركزي، له صلة بفيروس كورونا البشري OC43 وفيروس كورونا البقري. فيرروس كورونا البشري HKU1 -كغيره من الفيروسات المذكورة سابقا- له أصول ترجع إلى القوارض كذلك.
إصابة البشر
| ميرس-كوف | سارس-كوف | سارس-كوف-2 | |
|---|---|---|---|
| المرض | ميرس | سارس | كوفيد 19 |
| التفشيات |
2012، 2015، 2018 |
2002-2004 | جائحة 2019-2021 |
| الوبائيات | |||
| تاريخ أول حالة محددة |
يونيو 2012 | نوفمبر 2002 | ديسمبر 2019 |
| مكان أول حالة محددة |
جدة، السعودية |
غوانغدونغ، الصين |
ووهان، الصين |
| متوسط العمر | 56 | 44 | 56 |
| نسبة الجنس (ذ:إ) | 3.3:1 | 0.8:1 | 1.6:1 |
| الحالات المؤكدة | 2494 | 8096 | 668٬843٬945 |
| الوفيات | 858 | 774 | 6٬739٬355 |
| نسبة الحالات المتوفية | 37% | 9.2% | 1٫01% |
| الأعراض | |||
| الحمى | 98% | 99–100% | 87.9% |
| السعال الجاف | 47% | 29–75% | 67.7% |
| ضيق النفس | 72% | 40–42% | 18.6% |
| الإسهال | 26% | 20–25% | 3.7% |
| التهاب الحلق | 21% | 13–25% | 13.9% |
| استخدام التنفس الاصطناعي |
24.5% | 14–20% | 4.1% |
|
ملاحظات | |||
تختلف فيروسات كورونا اختلافا ملحوظا في عامل الخطر حيث يمكن لبعضها أن يقتل أكثر من 30٪ من المصابين مثل ميرس-كوف، والبعض الآخر غير ضار نسبيا ويسبب زكاما خفيفا. يمكن أن تسبب فيروسات كورونا زكاما مصحوبا بأعراض حادة مثل: مثل الحمى والتهاب الحلق بسبب الزوائد الأنفية المنتفخة. يمكن أن تسبب فيروسات كورونا ذات الرئة (إما ذات الرئة الفيروسي المباشر أو ذات رئة بكتيري ثانوي) والتهاب القصبات (إما التهاب قصبات فيروسي مباشر أو التهاب قصبات بكتيري ثانوي). فيروس كورونا البشري المكتشف عام 2003 -سارس-كوف- الذي يسبب متلازمة تنفسية حادة وخيمة (سارس) ، له إمراضية فريدة لأنه يسبب التهابات الجهاز التنفسي العلوي والسفلي.
توجد ستة أنواع معروفة من فيروسات كورونا البشرية، وينقسم نوع واحد إلى سلالتين مختلفتين، ما يجعل العدد الكلي سبع سلالات يمكنها إصابة البشر. أربع سلالات من بين هذه السلالات السبع تتسبب في أعراض خفيفة عموما، ورغم تكيف البشر معها، ربما كانت هذه الفيروسات أكثر حدة في الماضي:
- فيروس كورونا البشري OC43 (HCoV-OC43).
- فيروس كورونا البشري HKU1 (HCoV-HKU1).
- فيروس كورونا البشري 229E (HCoV-229E).
- فيروس كورونا البشري NL63 (HCoV-NL63).
ثلاث فيروسات كورونا يمكن أن تسبب أعراضا حادة:
- فيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة النوع 1 (سارس-كوف) ويسبب المتلازمة التنفسية الحادة الوخيمة (سارس).
- فيروس كورونا المرتبط بمتلازمة الشرق الأوسط التنفسية (ميرس-كوف) ويسبب متلازمة الشرق الأوسط التنفسية (ميرس).
- فيروس كورونا المرتبط بالمتلازمة التنفسية الحادة الشديدة النوع 2 (سارس-كوف-2) ويسبب مرض فيروس كورونا 2019 (كوفيد 19).
الزكام
يمكن أن تتسبب فيروسات كورونا البشرية (229E ،OC43 ،NL63 ،HKU1) في الإصابة بالزكام، وتنتشر هذه الفيروسات الأربعة على نطاق واسع بين السكان، وتتسبب في ظهور أعراض خفيفة. تشير الإحصائيات إلى أن 15٪ من حالات الزكام تسببها هذه الفيروسات الأربعة (وتشير بيانات أخرى إلى 20٪-30٪ ) في حين أن 50% من الحالات يسببها الفيروس الأنفي. في المناطق ذات المناخ المعتدل، تحدث الإصابة بهذه الفيروسات غالبا في فصل الشتاء، ولا يوجد فرق كبير في أوقات الإصابة في مناطق المناخ المداري. قد تتنافس فيروسات كورونا الأربعة مع بعضها البعض ويكون انتشارها بين السكان دوريا. مع أن فيروسات كورونا هذه تسبب الزكام فقط، فقد افتُرض أنها ربما تسببت في أوبئة حادة عندما ظهرت لأول مرة منذ قرون أو عقود مضت، ثم تطورت بمرور الوقت وتأقلم عليها البشر وأصبحت هذه السلالات أقل إمراضا. مثل فيروس كورونا البشري OC43 وفيروس كورونا البقري اللذان تشعبا عن النوع فيروس كورونا بيتا 1 في العقد 1890، حيث انتقل الفيروس البقري عبر الأنواع واكتسب القدرة على إصابة البشر، وعلى هذا الأساس افترض بعض العلماء أن جائحة الإنفلونزا (1889–1890) قد يكون سببها فيروس OC43 الناتج من هذا الانتقال عبر الأنواع وليس فيروس الأنفلونزا.
المتلازمة التنفسية الحادة الوخيمة (سارس)

في نهاية عام 2002 ظهرت ذات رئة غير نموذجية في مقاطعة غوانغدونغ بالصين، وتطورت لاحقا إلى تفشي السارس 2002-2004 الذي استمر حوالي عام تقريبا. انتشر الوباء إلى 29 دولة وأصاب أزيد من 8000 شخص توفي منهم 774 فردا. في مارس 2003 سمت منظمة الصحة العالمية بشكل رسمي المرض باسم السارس، ووجد العديد من العلماء أن الوباء سببه أحد فيروس كورونا وكان أول فيروس كورونا يسبب مرضا خطيرا. أرجع العلماء مصدر الفيروس المباشر إلى زباد النخل الذي بيع في الأسواق، وكان تماثل التسلسل بين فيروس سارس-كوف زباد النخل وفيروس السارس البشري 99.8%، لكن المستودع الطبيعي لهذا الفيروس يُفترض أن يكون أحد الخفافيش مثل: خفاش حدوة الحصان الصيني الأصهب أو خفاش الحذوة الكبير.فترة حضانة فيروس السارس حوالي 4 إلى 6 أيام وبعدها تظهر أعراض مماثلة للإنفلونزا والالتهاب الرئوي. في الحالات الحادة يمكن أن يعاني المرضى من صعوبات في التنفس وقد يتطور الأمر إلى متلازمة الضائقة التنفسية الحادة (ARDS). تصبح أعراض الإصابة أكثر حدة بعد أن يبدأ جهاز المناعة في محاربة الفيروس حيث تبين أن بعض الأعراض يسببها فرط الاستجابة المناعية الخاصة بالمريض. بعض مرضى السارس مازالوا يعانون من العديد من المضاعفات مثل: التليف الرئوي، هشاشة العظام والنخر اللاوعائي بعد الشفاء، وتطور لدى مرضى آخرين اضطراب الكرب التالي للصدمة النفسية أو اضطراب مزاجي.
متلازمة الشرق الأوسط التنفسية (ميرس)

في 2012 ظهر فيروس كورونا جديد سُمي لاحقا بفيروس كورونا متلازمة الشرق الأوسط التنفسية (ميرس-كوف)، حيث تم عزله من رئة مريض مصاب بأعراض تنفسية حادة في المملكة العربية السعودية. ويُعتقد أنه انتقل إلى البشر من الجمال، حيث وجدت دراسة أن فيروس كورونا الذي أصاب الجمال العربية البالغة وصغارها مطابق بنسبة 99.9% لجينومات ميرس-كوف البشرية. أما أصل هذا الفيروس ومستودعه الطبيعي فيُعتقد أنه يرجع إلى الخفافيش، وأن له صلة وثيقة بفيروس كورونا خفاش تيلونيكترايس HKU4 وفيروس كورونا خفاش بيبيسترالوس HKU5. اعتبارا من يناير 2021، أُبلغ عن 2500 حالة منها 45 حالة في 2020. وتوفي حوالي 35% من الأفراد الذين أصيبوا بالفيروس. وفضلا عن التفشي الأول عام 2012 حدثت تفشيات أخرى أحدها في كوريا الجنوبية عام 2015 وآخر في السعودية عام 2018.فترة حضانة فيروس ميرس حوالي 5.5 يوم، ويسبب أعراضا منها الحمى، السعال، الإسهال وضيق التنفس. بعض المرضى لا تظهر عليهم الأعراض، والحالات الخطيرة يمكن أن تسبب ذات رئة وخيمة ومتلازمة الضائقة التنفسية الحادة. الفيروس معدي نسبيا، وفي الغالب يصيب الأفراد الذين هم في اتصال مباشر مع المرضى، ولا ينتقل غالبا خارج المستشفى.
مرض فيروس كورونا 2019 (كوفيد 19)

في ديسمبر 2019، حدث تفشي التهاب رئوي جديد في مدينة ووهان بمقاطعة خوبي بالصين. ووجد العلماء في 31 ديسمبر أن التفشي سببه سلالة جديدة من فيروسات كورونا، والتي سميت لاحقا سارس-كوف-2 بواسطة اللجنة الدولية لتصنيف الفيروسات. يملك سارس-كوف-2 تماثل تسلسل قدره 79% مع سارس-كوف. ويُعتقد أن الخفافيش هي المستودع الطبيعي الأرجح له، حيث يملك فيروس كورونا الخفافيش RaTG13 تسلسلا متماثلا بنسبة 96.1% والفيروس الجديد المكتشف في اللاوس BANAL-52 نسبة 96.8%، توحي نسبة التماثل بين هذه الفيروسات الخفاشية وسارس-كوف-2 بأن البشر قد أصيبوا بالفيروس عبر مضيف وسيط، لكن بقي هذا الوسيط وهو المصدر المباشر لإصابة البشر غير معروف.
اعتبارا من 22 يناير 2023 يوجد على الأقل 6٬739٬355 حالة وفاة مؤكدة وأزيد من 668٬843٬945 إصابة مؤكدة بسبب جائحة كوفيد 19، التي كان لها تأثير كبير على المجتمع والإقتصاد العالمي وسببت أسوء ركود اقتصادي منذ الكساد الكبير في عقد 1930. أعراض كوفيد 19 متنوعة وتختلف من فرد لآخر، ومدة حضانة فيروس سارس-كوف-2 تتراوح بين 1 إلى 14 يوما. على الأقل ثلث الأفراد المصابين لا تظهر عليهم الأعراض، لكن بإمكانهم نقل الفيروس، معظم أعراض كوفيد 19 خفيفة أو متوسطة ومن بينها: الحمى، السعال، الإعياء، فقدان الشم المؤقت وذات الرئة الخفيفة. تعاني نسبة صغيرة من المرضى من أعراض حادة منها: ضيق التنفس ونقص التأكسج، أما الحالات الخطرة فقد تعاني من فشل تنفسي، صدمة وفشل الأعضاء المتعدد. بعض المرضى تبقى لديهم أعضاء متضررة بعد الشفاء تفشل في التعافي التام، ويعانون من مضاعفات ما بعد الإصابة لأمد طويل.
إصابة الحيوانات
عُرِف أن فيروسات كورونا تسبب حالات مرضية في الطب البيطري منذ ثلاثينيات القرن الماضي، وهي تصيب مجموعة متنوعة من الحيوانات منها: الخنازير والأبقار والخيول والجمال والقطط والكلاب والقوارض والطيور والخفافيش. تصيب معظم فيروسات كورونا المرتبطة بالحيوانات السبيل الهضمي بواسطة الطريق الفموي الشرجي. تم تركيز جهود بحثية معتبرة لمعرفة الإمراضية الفيروسية الخاصة بفيروسات كورونا الحيوانية هذه، وخاصة من قبل علماء الفيروسات المهتمين بالأمراض البيطرية والحيوانية.
الدواجن والماشية

يمكن لفيروسات كورونا أن تصيب أنواعا مختلفة من الدواجن والماشية. فيروس التهاب الشعب الهوائية المعدي (IBV) هو فيروس كورونا غاما يصيب الدواجن عبر الهباء الجوي أو الطريق الفموي الشرجي، ويسبب التهابا شعبيا معديا ويصيب الجهاز البولي التناسلي فضلا عن السبيل التنفسي. يملك فيروس التهاب الشعب المعدي معدل نفوق مرتفع، وانتشارا سريعا، ويؤثر على إنتاج البيض ويسبب خسائر اقتصادية كبيرة، وتوجد العديد من اللقاحات للوقاية منه. بعض السلالات الشبيهة بـIBV يمكنها أن تصيب الدجاج الرومي وتسبب التهاب الأمعاء منها فيروس كورونا الدجاج الرومي (TCV). يُصنف فيروس التهاب الشعب المعدي إلى جانب مختلف سلالات الدجاج الرومي وبعض الفيروسات التي تصيب الطيور البرية إلى نوعين: فيروس كورونا الطيري، وفيروس كورونا الطيري 9203 تحت جنس فيروسات كورونا غاما.
تصاب الماشية مثل الخنازير والأبقار والأحصنة بفيروسات كورونا كذلك. من بين فيروسات كورونا التي تصيب الخنازير: فيروس الإسهال الوبائي الخنزيري (PEDV)،فيروس التهاب المعدة والأمعاء الساري الخنزيري (TGEV)،فيروس كورونا دلتا الخنزيري (PdCV)، فيروس التهاب الدماغ والنخاع الشوكي الراصي الخنزيري (PHEV) وفيروس كورونا متلازمة الإسهال الحاد الخنزيري (SADS-CoV). باستثناء PHEV الذي يصيب النسيج العصبي كذلك، تصيب فيروسات كورونا الخنزيرية الجهاز الهضمي وتسبب القيء والإسهال وأعراضا أخرى. فيروسا TGEV وPHEV متنشران بين الخنازير لأزيد من 10 سنوات أما SADS-CoV وPdCV فهما فيروسان حديثا النشأة، ورغم أن PEDV متواجد منذ 1970 إلا أن سلالة شديدة الإمراض ظهرت في 2010 وسببت خسائر ملحوظة في تربية الخنازير في بلدان مختلفة. من بين الفيروسات الناشئة، تسلسل SADS-CoV مماثل جدا لفيروس كورونا خفاش حدوة الفرس الصيني HKU2، وتسلسل PEDV مماثل لفيروس كورونا خفاش إلف الظلام 512 وقد يرجع أصلهما إلى انتقال عبر الأنواع من الخفافيش. ربما يرجع أصل PdCV إلى فيروسات كورونا الطيرية فتسلسله متماثل جدا مع فيروس كورونا الدوري HKU17 وفيروس كورونا السمان HKU30 ويُفترض أنهما ينتميان إلى نفس النوع.
فيروس كورونا البقري الذي يصيب الأبقار قريب الصلة بفيروس كورونا البشري OC43، ويسبب أعراضا مثل الإسهال لدى العجول. يسبب فيروس كورونا الفرسي الذي يصيب الخيول الإسهال كذلك، ونادرا ما يكون قاتلا.
الحيوانات الأليفة

تصاب الحيوانات الأليفة مثل: القطط والكلاب وابن مقرض بفيروسات كورونا كذلك. ينتمي فيروس كورونا السنوري إلى فيروسات ألفا ويمكن أن يُقسم إلى نوعين: فيروس كورونا القططي المعوي (FECV) الذي يصيب الأمعاء ويسبب أعراضا خفيفة، وفيروس التهاب البريتون المعدي السنوري (FIPV) الذي يمكن أن يسبب التهاب البريتون المعدي السنوري الحاد. ينتقل FECV عبر الطريق الشرجي الفموي ويمكن أن يتطفر إلى FIPV بعد الإصابة، ثم ينتقل لإصابة مختلف أعضاء القطط. معدل النفوق بالتهاب البريتون المعدي السنوري مرتفع وعادة ما تموت القطط خلال عام واحد من ظهور الأعراض.
معروف بأن اثنين من فيروسات كورونا تصيب الكلاب: فيروس كورونا الكلبي (CCoV) وفيروس كورونا التنفسي الكلبي (CRCoV). الأول من فيروسات كورونا ألفا ويسبب أعراضا معدية معوية خفيفة، أما الثاني فهو من فيروسات كورونا بيتا ويصيب الجهاز التنفسي وهو أحد الممرضات التي تسبب سعال الكلاب. يوجد كذلك نوعان من فيرووسات كورونا التي تصيب ابن مقرض:فيروس كورونا ابن مقرض ويسبب التهاب الأمعاء، وفيروس كورونا جهازي ابن مقرضي (FSC) وهو أشد خطرا ويسبب مرض جهازي مماثل لالتهاب البريتون المعدي السنوري.
الحيوانات المختبرية
يمكن لفيروسات كورونا أن تصيب العديد من الحيوانات المستخدمة في المختبرات. فيروس التهاب الكبد الفأري (MHV) كان أكثر فيروس كورونا تمت دراسته حيويا ومخبريا وعلى المستوى الجزيئي قبل اكتشاف سارس-كوف، ويصيب صغار فئران التجارب مسببا معدلات نفوق عالية، بينما يسبب أعراضا خفيفة قابلة للعكس لدى الفئران البالغة، لكنه يثير استجابة مناعية يمكن أن تؤثر على تفسير النتائج التجريبية. ينقسم MHV إلى عدة سلالات وتتنوع الأنسجة التي يصيبها وحدة الأعراض التي يسببها. يسبب MHV-1 أعراضا تنفسية وتسبب السلالتان A59 وMHV-3 التهاب الكبد ويسبب JHV التهاب الدماغ. يحفز الفيروس الأخير استجابات مناعية لدى الفئران تسبب أضرارا لأغمدة الميالين في العصبونات، لذلك يُستخدم كنموذج حيواني لدراسة التصلب المتعدد. من فيروسات كورونا التي تصيب الجرذان: فيروس التهاب الغدد اللعابية والدمعية (SDAV) وفيروس كورونا باركر الجرذي (RCoV-P)، يمكن أن تصيب السلالة الأولى الأعين والغدد الدمعية والغدد اللعابية ويمكن كذلك أن تؤثر على تكاثر الجرذان وتسبب تغيرات سلوكية وهي أحد أهم الممرضات لجرذان التجارب. تصنف هاتان السلالتان الجرذيتان مع مختلف سلالات MHV الفأرية كنوع واحد ويشار إليهم جميعا باسم فيروس كورونا الموريني.
يوجد فيروس كورونا يمكن أن يصيب الأرانب ويسبب التهاب الأمعاء مع معدلات نفوق عالية، ويسمى فيروس كورونا المعوي الأرنبي (RECV) وينتقل عبر الطريق الشرجي الفموي. فضلا عن ذلك، تُصاب الأرانب المستخدمة لزراعة اللولبية الشاحبة بممرضات غير معروفة يفترض أن تكون فيروس كورونا، وتوجد حالات قليلة فقط لإصابة الأرانب بهذين النوعين من فيروس كورونا. في 2012، وجد بعض الباحثين فيروس كورونا الأرنبي HKU14 في الأرانب التي بيعت في الأسواق، وهو فيروس بيتا يصيب الأرانب من دون أعراض ظاهرة. توجد كذلك تقارير حول إصابة فيروس كورونا للكابياء الخنزيرية وتسببه في أعراضٍ مثل الإسهال والتهاب الأمعاء، لكن مدى انتشار الفيروس في أوساط الكابياء الخنزيرية وعلاقته بفيروسات كورونا الأخرى مازالت غير معروفة.
الحيوانات البرية
زيادة على الحيوانات سالفة الذكر التي يربيها الإنسان، توجد العديد من سلالات فيروس كورونا التي تصيب الطيور والثدييات البرية. بعد حدث السارس في عام 2003، اكتُشِف عدد كبير من فيروسات كورونا ألفا وبيتا في مجموعة متنوعة من الخفافيش البرية، ويرتبط العديد منها ارتباطا وثيقا بسلالات سارس-كوف وميرس-كوف وسارس-كوف-2 التي تسببت في التفشيات. وُجدت العديد من سلالات هذين الجنسين في ثدييات برية صغيرة مثل: القوارض وأرنبيات الشكل والقنافذ والزبابات. فضلا عن ذلك، وُجد في مجترات مثل: الرنة والغزلان والبيسون والظباء والزرافات سلالات فيروس كورونا مماثلة لفيروس كورونا البقري، ووجد كذلك لدى الثدييات البحرية مثل الفقميات والحيتانيات فيروسات من الجنس ألفا وغاما على التوالي (فيروس كورونا الحوتي) أما لدى الطيور البرية فقد عُثر على سلالات كثيرة من غاما ودلتا، وصُنف العديد من سلالات غاما تحت النوع الجديد فيروس كورونا البطي 2714. معظم الأنواع في جنس دلتا تصيب الطيور البرية. بالإضافة إلى الثدييات والطيور، وُجدت سلالةٌ لفيروس كورونا تصيب الزواحف (السحالي الصينية).
الوقاية

بين الأمراض البشرية، لا توجد لقاحات لفيروسات كورونا البشرية الأربعة التي تسبب الزكام، وسبيل تجنب الإصابة بها هو غسل الأيادي وتجنب لمس الأعين والأنف والفم وارتداء الكمامات. اعتبارا من 2021، مازال لا يوجد لقاح فعال للوقاية من السارس، وميرس، لكن بعض اللقاحات ضد ميرس هي قيد التطوير. السارس مرض ينتقل عبر الرذاذ التنفسي ويمكن لقناع N95 منع الإصابة به بفعالية، وبسبب قوة الإعداء الكبيرة لمرضى الحالات الحادة فإن العزل والحجر الصحي إجراء فعال من أجل الحد من انتشاره، والمرضى الذين يتم عزلهم قبل اليوم الخامس من الإصابة نادرا ما يقومون بنقل الفيروس للآخرين.
فضلا عن الحفاظ على النظافة الشخصية وعزل المرضى، من إجراءات منع انتشار كوفيد 19: الحرص على التباعد الاجتماعي، إلغاء التجمعات غير الضرورية، العمل والدراسة عبر الإنترنيت ومنع انتشار الجائحة عبر تقييد السفر. طُورت العديد من اللقاحات لكوفيد 19 وتنصح منظمة الصحة العالمية الأفراد بأخذ جميع الجرعات الموصى بها بواسطة الهيئات الصحية التي يتبعون لها بمجرد أن يحين دورهم بما في ذلك جرعة معززة إذا تمت التوصية بها.
لدى الحيوانات، تطوير اللقاحات ضد فيروسات كورونا صعب لأنها عرضة للتأشيب الجيني، ويمكن للقاحات أن تعزز حدوث طفرات أخرى. لا يمكن الوقاية من التهاب البريتون المعدي السنوري الذي تسببه الإصابة بفيروس كورونا السنوري باللقاحات، واللقاحات المطورة ضده في الماضي لم تفشل في الوقاية منه وحسب بل تسببت في تفاقم الأعراض. حاليا، توجد لقاحات يمكن أن تقي من فيروس التهاب الشعب الهوائية المعدي (IBV)، فيروس التهاب المعدة والأمعاء الساري (TGEV)، وفيروس الإسهال الوبائي الخنزيري (PEDV) وفيروس كورونا الكلبي، لكن تأثيرها الوقائي غير تام ويمكن أن تسبب تأشيبا جينيا لتوليد سلالات جديدة، خضع فيروس التهاب المعدة والأمعاء الساري إلى طفرة حذف تسلسل ونتجت سلالة متطفرة تسمى فيروس كورونا التنفسي الخنزيري (PRCV) تسبب أعراضا خفيفة ونجح التلقيح بهذه السلالة الموهنة من الوقاية ضد تفشيات جديدة حادة لـTGEV خلال الـ30 عاما الماضية.
ملاحظات
انظر أيضًا
المراجع
باللغة الإنجليزية
باللغة العربيَّة
مراجع إضافية
- Alwan A، Mahjour J، Memish ZA (2013). "Novel coronavirus infection: time to stay ahead of the curve". Eastern Mediterranean Health Journal. 19 Suppl 1: S3–4. PMID 23888787. مؤرشف من الأصل في 2020-03-24.
- Laude H، Rasschaert D، Delmas B، Godet M، Gelfi J، Charley B (يونيو 1990). "Molecular biology of transmissible gastroenteritis virus". Veterinary Microbiology. ج. 23 ع. 1–4: 147–54. doi:10.1016/0378-1135(90)90144-K. PMID 2169670.
- Sola I، Alonso S، Zúñiga S، Balasch M، Plana-Durán J، Enjuanes L (أبريل 2003). "Engineering the transmissible gastroenteritis virus genome as an expression vector inducing lactogenic immunity". Journal of Virology. ج. 77 ع. 7: 4357–69. doi:10.1128/JVI.77.7.4357-4369.2003. PMC 150661. PMID 12634392.
- Tajima M (1970). "Morphology of transmissible gastroenteritis virus of pigs. A possible member of coronaviruses. Brief report". Archiv für die Gesamte Virusforschung. ج. 29 ع. 1: 105–8. doi:10.1007/BF01253886. PMID 4195092.